

2. Энергетика транспортных процессов у микроорганизмов
 Источником
энергии, обеспечивающим активный
транспорт в клетки различных
микроорганизмов, в большинстве случаев
является трансмембранный электрохимический
потенциал ионов водорода, который может
создаваться за счет переноса электронов
или распада АТФ под влиянием мембранной
АТФазы. Переносчики, имеющие места
связывания протонов и молекул субстрата,
используют Н+-
электрохимический мембранный потенциал
(р
- протонодвижущую силу)
для транспорта в клетку ионов водорода
и питательных веществ.
Источником
энергии, обеспечивающим активный
транспорт в клетки различных
микроорганизмов, в большинстве случаев
является трансмембранный электрохимический
потенциал ионов водорода, который может
создаваться за счет переноса электронов
или распада АТФ под влиянием мембранной
АТФазы. Переносчики, имеющие места
связывания протонов и молекул субстрата,
используют Н+-
электрохимический мембранный потенциал
(р
- протонодвижущую силу)
для транспорта в клетку ионов водорода
и питательных веществ.
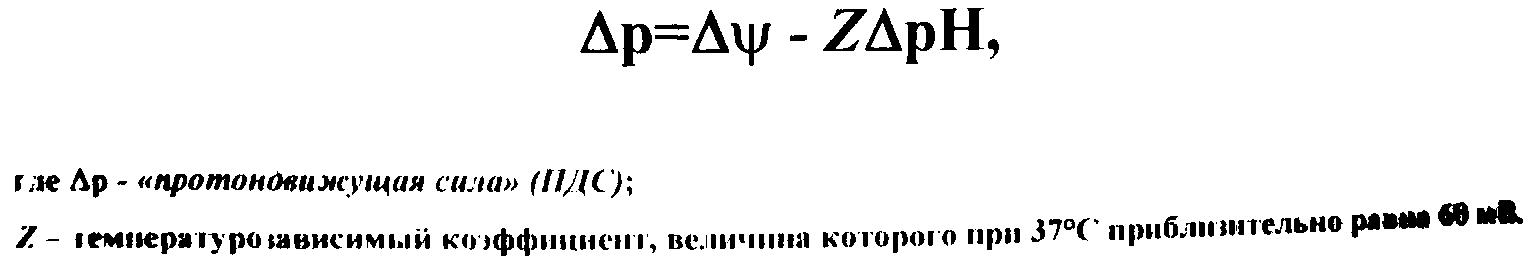
Связывание с протоном повышает сродство переносчика к субстрату, а высвобождение его от протона на внутренней поверхности мембраны - понижает это сродство. Такой совместный транспорт одним переносчиком двух субстратов в одном направлении называется симпортом. Унипорт - переносчик транспортирует только один субстрат. Многие питательные вещества поступают в клетки микробов также за счет симпорта с ионами Na+ или К+. Существует еще механизм антипорта, когда один переносчик транспортирует два субстрата, но в противоположном направлении.
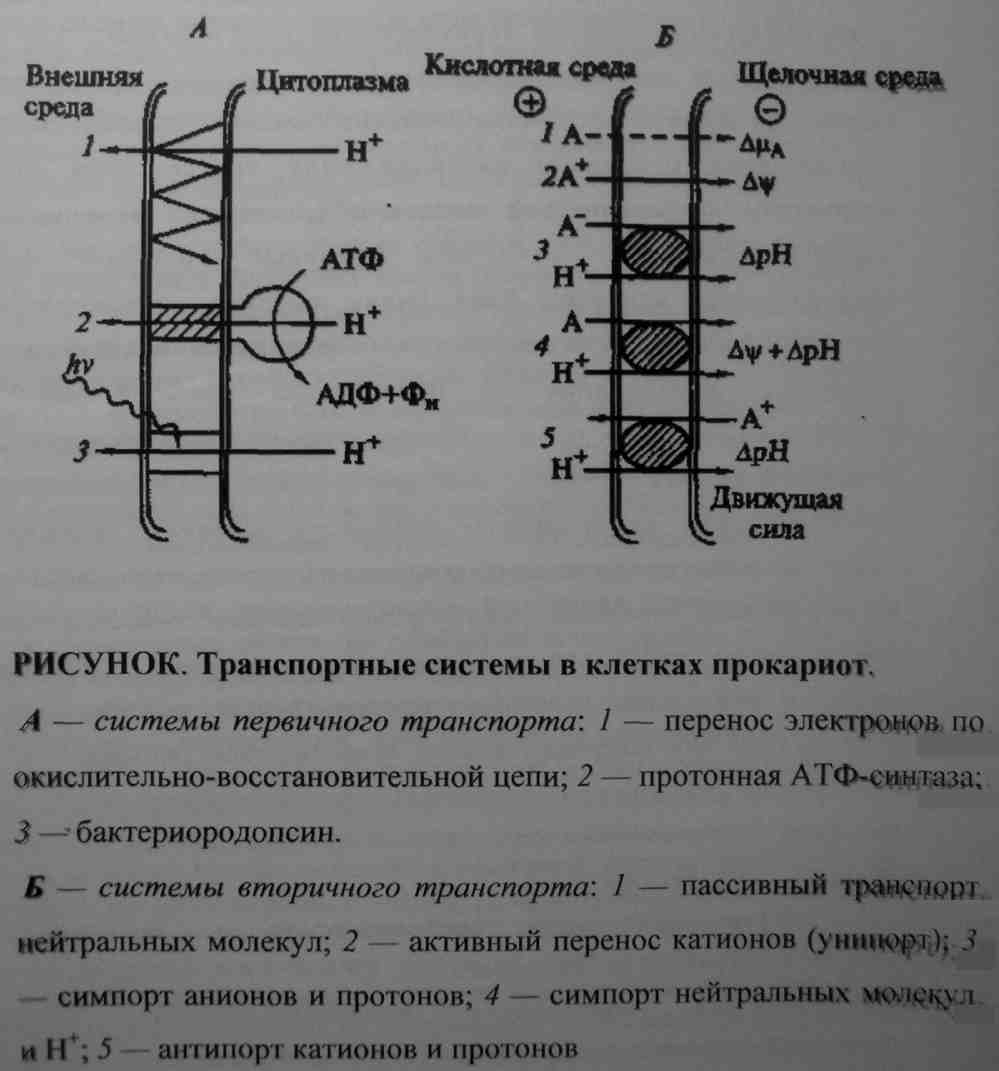
У микроорганизмов, в частности у Е. coli обнаружены также системы активного транспорта, которые используют химическую энергию АТФ. Такие системы обычно функционируют с помощью расположенных в периплазме связывающих белков. Это водорастворимые белки, обладающие высоким сродством к некоторым аминокислотам, витаминам, пептидам, сахарам и органическим кислотам. Сами они не могут транспортировать субстраты через плазматическую мембрану, но способны стимулировать активность мембранных компонентов системы транспорт. Физиологическая роль связывающих белков состоит в том, чтобы концентрировать в переплазматическом пространстве определенные соединения, имеющиеся в среде, а также в том, чтобы препятствовать выходу в среду питательных веществ, вытекающих из цитоплазмы.
Существует два основных пути, по которым может тратиться энергии на транспортный процесс.
1. Энергия затрачивается на химическую модификацию субстрата в процессе транслокации, что делает его не способным ни взаимодействовать с переносчиком, ни проникать через мембрану диффузионным путем. По этой причине продукт модификации субстрата накапливается в клетке. Следовательно, транспорт вещества непосредственно связан с первыми этапами его метаболизма. Транспортные системы такого типа получили название систем «переноса радикалов» или «векторного метаболизма». У микроорганизмов наиболее изучены три типа таких систем.
Примером одной из систем переноса групп является фосфотрансферазная система (ФТС). Она транспортирует многие углеводы и их производные, поступающие в клетку в виде фосфатных эфиров (фосфоуглеводы). При транспорте углеводов с помощью механизма переноса групп в клетке сразу же оказывается фосфорилированное производное.
2. Энергия затрачивается на химическую модификацию переносчика в процессе транслокации, что затрудняет или делает невозможной обратную транслокацию субстрата из клетки. В этом случае источником энергии для транспортных систем, содержащих связывающие белки (первичный активный транспорт), может служить АТФ либо трансмембранный электрохимический потенциал.