

Глава VI половинные и целые зародыши
В классических опытах Вильгельма Ру (1883), вокруг которых в течение многих лет происходила большая дискуссия, один из первых двух бластомеров яйца лягушки повреждался раскаленной иглой. Другой же бластомер развивался и образовывал половину эмбриона. Ру сделал из своих опытов вполне естественный вывод, что части зародыша закладываются, начиная уже с первого деления, путем диференциальных дроблений. Он пошел даже дальше и приписал источник этой диференциации качественному распределению хроматинового вещества. Позднее Вейсман на этом же самом допущении основал всю свою теорию процесса развития. Однако даже в те времена имелось уже много данных в пользу того, что, как при первых, так и при всех последующих делениях яйца, каждая хромосома продольно расщепляется на две в точности равные части. Это последнее заключение полностью подтвердилось открытыми впоследствии генетическими фактами, которые по существу доказывают не только видимое деление хромосом, но также невидимое деление самих генов.
Несколькими годами позже Ганс Дриш (1891) экспериментировал с яйцами морского ежа и обнаружил, что если разделить первые две клетки, то каждая из них развивается в целый эмбрион половинной величины. Этот результат находился в резком противоречии с полученным Ру и приводил к противоположному выводу, а именно, что при первых делениях яйца диференцировка не имеет места. По выражению Дриша, каждая из первых двух клеток является «тотипотентной». Однако эти два опыта Дриша и Ру были неидентичными. В яйце лягушки поврежденный бластомер, остававшийся еще живым, соприкасался со своим партнером, тогда как в яйце морского ежа два {74} бластомера полностью разделялись. Это различие, однако, не должно сильно влиять на сделанный Ру вывод, так как, если диференцировка начинается с первого деления, то это должно относиться к обоим случаям.
Со времени этих первых опытов и их интерпретации было проведено много других сходных экспериментов не только с яйцами лягушки и морского ежа, но и с яйцами многих других видов. Прежде чем перейти к обсуждению относительной значимости споров, которые были основаны на этих наблюдениях Ру и Дриша, необходимо рассмотреть последние опыты.
Опыты с яйцами морского ежа
Когда первые два бластомера морского ежа отделяются друг от друга, каждый из них округляется. При следующем делении каждый бластомер делится точно так же, как он
|
|
|
Рис. 40. а, b, с— четырех-, восьми- и шестнадцатиклеточная стадии яйца морского ежа; а1,b1,с1— первые три дробления изолированной половины бластомера (по Дришу). |
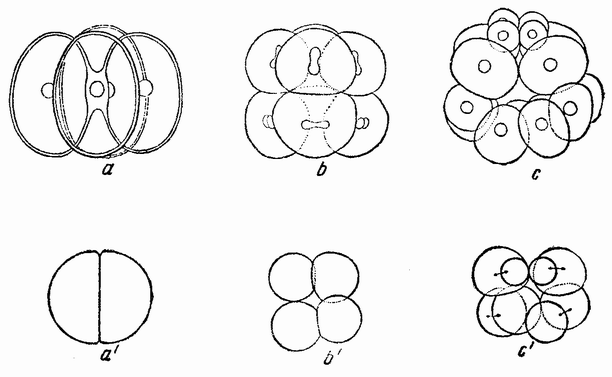
делился бы, если бы он оставался соединенным со своим соседом, т.е. борозда проходит через анимальный полюс (рис.40, a1). Следующее деление является экваториальньш, а следующее за ним образует у вегетативного полюса микромеры (рис.40,b1—с1). При первых делениях открытая сторона {75} половинной бластулы закрывается и, по мере того как продолжаются деления, внутри бластулы появляется пространство, заполненное жидкостью. Затем происходит га-
|
|
|
Рис. 41. Целые и половинные гаструлы и плутеусы морского ежа; последние развились из половинных бластомеров. |

струляция, после чего образуется типичный плутеус (рис. 41, b—b1).
Способ закрытия бластулы и инвагинации первичного кишечника представляет в некоторых отношениях {76} теоретический интерес. Эти процессы могут быть изучены путем окраски вегетативной области половины шестнадцатиклеточной стадии. Полученные результаты показывают, что открытая сторона половины бластулы закрывается и клетки анимального и вегетативного полюсов (равно как и клетки с боков бластулы) сближаются. Инвагинированный первичный кишечник начинается в середине энтодермального поля. Эти отношения показаны на рис. 42, на котором изображена целая бластула (b), ее нормальный способ
|
|
|
Рис. 42. b— нормальная бластула и способ ее инвагинации (а),с— для сравнения инвагинация половинной бластулы (по Герстадиусу). |
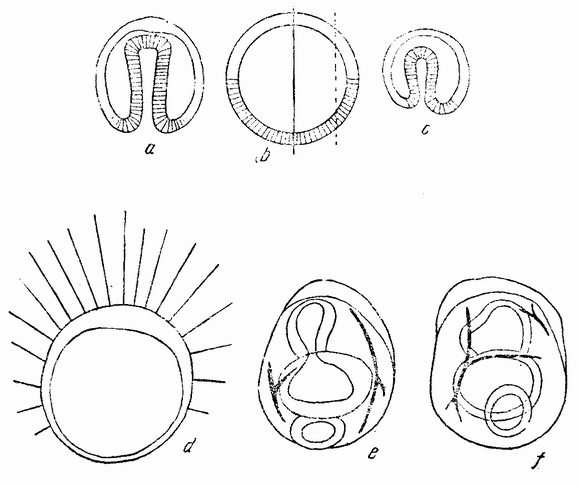
гаструляции (а) и гаструляция половинной гаструлы (с). Инвагинация происходит симметрично, причем впячивается только энтодермальное вещество. Это, казалось бы, означает, что первоначальный центр энтодермального вещества (которое после разделения лежит на стороне половинной бластулы) не является центром гаструляции половинной бластулы, а что этот центр восстанавливается в середине энтодермального вещества. Это смещение центра можно представить следующим образом. Клетки на анимальном полюсе приходят в соприкосновение с клетками {77} вегетативного полюса, где сначала был центр энтодермы. Если затем, благодаря этому соприкосновению, энтодермальные клетки теряют некоторые из своих способных к диффузии веществ, и, наоборот, эктодермальные клетки отдают энтодерме некоторые из своих веществ, то наиболее концентрированное вещество энтодермы окажется лежащим ближе к середине энтодермальной массы. Образуется новый центр, который становится центром гаструляции. Если на четырехклеточной стадии яйца морского ежа изолировано четыре бластомера, то каждый сегмент является частью целого. При втором делении образуется только единственный микромер. Развивается бластула вчетверо меньшей величины, которая гаструлируется и превращается в эмбрион-плутеус, величина которого тоже вчетверо меньше. Правда, не было обнаружено, что все четыре бластомера одного и того же яйца ведут себя одинаковым образом, но надо думать, что это так.
Надо отметить, что как половинки, так и четвертушки бластомеров содержат все вещества яйца от анимального до вегетативного полюсов, но после третьего деления четыре верхние клетки оказываются содержащими только анимальное вещество, а нижние клетки только вегетативное. В действительности было обнаружено, что некоторые из изолированных восьмушек бластомеров гаструлируют, тогда как другие остаются плавающими бластулами. Однако потенции верхних и нижних клеток лучше всего видны при отделении четырех верхних бластомеров от четырех нижних. Было найдено, что четыре верхние (анимальные) клетки дают начало плавающей бластуле (рис. 42, d), которая не гаструлирует, но у которой может произойти инвагинация рта. Четыре нижних клетки дают начало гаструле, у которой впоследствии развивается пищеварительный тракт, разделенный на характерные три отдела. Может происходить и начало образования скелета, но рот все же не образуется (рис. 42,е—f). Из этого очевидно, что верхняя и нижняя половины развиваются только в частичные структуры, сходные с теми частями, которые развиваются из этих же самых участков целой бластулы.