

medvir_xxvii_2_2013
.pdf
Главный редактор
МихайловМ.И., чл.-корр.РАМН,профессор,д.м.н.
ЗАМЕСТИТЕЛЬ ГЛАВНОГО РЕДАКТОРА
НоводержкинаЮ.К.,д.м.н.
Редакционная коллегия
ГмыльА.П.,к.б.н. ГордейчукИ.В.,к.м.н. ДзагуроваТ.К.,к.м.н. J.Drobeniuc,Ph.D.
ИльченкоЛ.Ю.,д.м.н.,профессор
S.Kamili,Ph.D.
КаргановаГ.Г.,д.б.н. B.Klempa,Ph.D.
КозловскаяЛ.И.,к.б.н. D.H.Kruger,M.D.,Ph.D.
КюрегянК.К.,д.м.н. ЛукашевА.Н.,д.м.н. МатросовичМ.Н.,Ph.D.
ПогодинаВ.В.,д.м.н.,профессор РомановаЛ.Ю.,к.б.н.
D.Růžek,Ph.D.
ТкаченкоЕ.А.,д.м.н.,профессор ЧумаковК.М.,Ph.D.
Адрес редакции
142782г.Москва,
поселениеМосковский, поселокИнститутаполиомиелита, 27кмКиевскогошоссе Телефон:(495)841-90-07 Факс:(495)841-93-30
www.poliomyelit.ru
Данные об издательстве
ЗАО«ИздательствоИКАР».
Москва,117485,ул.АкадемикаВолгина,д.6.
Тел.:8(495)936-83-28,978-35-99. Тел./факс:8(495)330-89-77 www.ikar-publisher.ru
На обложке:
ТОМ XXVII (2) |
|
VOL XXVII (2) |
|
|
|
Editor-in-Chief
MikhailovM.I.,
CorrespondingmemberofRAMS,Professor,M.D.,Ph.D.,Sc.D.
DEPUTYCHIEFEDITOR
Y.K.Novoderzhkina,M.D.,Ph.D.,Sc.D.
Editorial Board
A.P.Gmyl’,Ph.D.
I.V.Gordeychuk,M.D.,Ph.D.
T.K.Dzagurova,M.D.,Ph.D.
J.Drobeniuc,Ph.D.
L.Yu.Ilchenko,Professor,M.D.,Ph.D.,Sc.D.
S.Kamili,Ph.D.
G.G.Karganova,Ph.D.,Sc.D.
B.Klempa,Ph.D.
L.I.Kozlovskaya,Ph.D.
D.H.Kruger,M.D.,Ph.D.
K.K.Kyuregyan,M.D.,Ph.D.,Sc.D.
A.N.Lukashev,M.D.,Ph.D.,Sc.D.
M.N.Matrosovich,Ph.D.
V.V.Pogodina,Professor,M.D.,Ph.D.,Sc.D.
L.Iu.Romanova,Ph.D.
D.Růžek,Ph.D.
E.A.Tkachenko,Professor,M.D.,Ph.D.,Sc.D.
K.M.Chumakov,Ph.D.
Editorial Office Address
Institutpoliomyelita, 27kmKievskoehighway, Moscow,142782 Russia
Phone:(495)841-90-07
Fax:(495)841-93-30 www.poliomyelit.ru
Publisher
IKARPublisher.
6,AkademikaVolginastr.,Moscow,117485,Russia. Ph.:8(495)936-83-28,978-35-99.
Fax:8(495)330-89-77 www.ikar-publisher.ru
COVER LEGEND:
Коинфекция гепатоцита вирусом TT-like (в ядре) и вирусом гепатита В (в цитоплазме) у больного с хроническим гепатитом. Электронная микрофотография, увеличе-
ниеХ15000
Co-infection of a hepatocyte with TT-like virus (in the nucleus) and hepatitis B (in the cytoplasm)inachronichepatitispatient.Electronmicrograph,magnificationХ15000

Содержание
Аденовирусный кератоконъюнктивит: возбудитель, эпидемиология, лечение
А.Н.Лукашев
ТерапияхроническогогепатитаС:настоящееи будущее
Н.И.Громова
Эпизоотологические характеристики геморрагической лихорадки с почечным синдромом на территории Тамбовской области
Е.С.Мутных,Е.В.Калинкина,Т.К.Дзагурова, А.Д.Бернштейн,Н.А.Коротина,Г.А.Толстова, Г.Д.Тихонова,С.Е.Соцкова,Е.А.Ткаченко
Нозоареал и структура заболеваемости геморрагической лихорадки с почечным синдромом на территории Тамбовской области
Е.С.Мутных,Т.К.Дзагурова,Е.В.Калинкина, А.Д.Бернштейн,Н.А.Коротина,Г.А.Толстова, Г.Д.Тихонова,С.Е.Соцкова,Е.А.Ткаченко
Анализ молекулярной динамики белков Е вариантов ВКЭ на основе разности корреляционных матриц
Л.И.Козловская,Д.И.Осолодкин,Г.Г.Карганова
Проблемы морфологической диагностики хронического гепатита С
Л.Ю.Ильченко,И.А.Морозов
Серологические маркеры инфицирования вирусами гепатитов среди условно здорового населения Республики Тыва
Т.В.Кожанова,Л.Ю.Ильченко,В.В.Клушкина, В.С.Кичатова,Н.В.Соболева, А.А.Сарыглар, И.В.Гордейчук,О.В.Исаева, К.К.Кюрегян, М.И.Михайлов
Правила представления рукописей авторами в журнал «Медицинская вирусология»
Порядок рецензирования рукописей научных статей, направляемых для открытого опубликования в редколлегию журнала «Медицинская вирусология»
Content
3Adenovirus keratoconjunctivitis: causative agent, epidemiology, clinical manifestations and treatment
A.N.Lukashev
16Therapy of chronic hepatitis c: current status and future prospects
N.I.Gromova
23Epizootological characteristics of natural foci of hemorrhagic fever with renal syndrome (HFRS) in Tambov region
E.S.Mutnykh,Е.V.Kalinkina,A.D.Bernshtein, T.K.Dzagurova,N.А.Korotina,G.A.Tolstova, G.D.Tikhonova,S.E.Sotskova,Е.А.Tkachenko
32Nozoareal and morbidity structure of hemorrhagic fever with renal syndrome in Tambov region
E.S.Mutnykh,T.K.Dzagurova,Е.V.Kalinkina, A.D.Bernshtein,N.А.Korotina,G.A.Tolstova, G.D.Tikhonova,S.E.Sotskova,Е.А.Tkachenko
42Analysis of molecular dynamics of TBEV variants’ E proteins based on difference of correlation matrices
L.I.Kozlovskaya,D.I.Osolodkin,G.G.Karganova
54Problems of morphological characteristics of chronic hepatitis C
I.A.Morozov,L.Yu.Ilichenko
74Serologic markers of infection with hepatitis viruses in conditionally healthy population of a region with high hepatitis morbidity
T.V.Kozhanova,L.Y.Ilchenko,V.V.Klushkina, V.S.Kichatova,N.V.Soboleva,A.A.Sargylar, I.V.Gordeychuk,V.V.Isaeva,E.Y.Malinnikova, K.K.Kyuregyan,M.I.Mikhailov
89/91 Author guidelines for journal «Medical virology»
94/95 Course of reviewing of the manuscripts
for publication in «Medical Virology»

medical virology. vol. XXVII (2) |
3 |
Аденовирусный кератоконъюнктивит: вирусологическая характеристика возбудителя, эпидемиология,лечение
А.Н. Лукашев
ФГБУ «Институт полиомиелита и вирусных энцефалитов имени М.П. Чумакова» РАМН
Резюме
Аденовирусыхорошоисследованыэкспериментально, но остаются относительно малоизученными с клинической точки зрения. Аденовирусы редко вызывают тяжелую патологию, но вносят заметный вклад в этиологию сравнительно легких заболеваний. В частности, аденовирусы являются одним из основных возбудителейконъюнктивита. Заболевание, как правило, заканчивается самопроизвольным выздоровлением без осложнений, но протекает 3–6 месяцев, а официально регистрируемая заболеваемость совершенно не отражает его социально-экономическую роль. На сегодняшнийденьнетметодовлеченияаденовирусного кератоконъюнктивита с доказанной эффективностью. В данном обзоре изложена молекулярнобиологическая и клиническая характеристика аденовирусов, а также эпидемиология, клиника и лечение аденовирусного кератоконъюнктивита.
Введение
Аденовирусы – ДНК-содержащие вирусы, способные инфицировать человека и других позвоночных. Аденовирусы были выделены в 1953 году из ткани носоглоточной миндалины (аденоидов), что и стало основанием для их названия. Изучение аденовирусов внесло большой вклад в развитие молекулярной биологии. Сплайсинг мРНК был открытв1975годуименноуаденовирусов.Аденовирусытакжебылипервоймодельюдляизучениявирусногоонкогенеза.Относительнаяпростотамодификации генома, культивирования и очистки вируса привела к тому, что аденовирусные векторы широко используются для генной терапии, уступая по популярности только лентивирусным векторам.
Классификация аденовирусов
Аденовирусы образуют семейство Adenoviridae. Насегодняшнийденьизвестноболее100аденовирусов, выделенных от разных позвоночных. Классификация по родам основана на хозяевах благодарястрогойспецифичностиаденовирусовотносительно вида-хозяина. Аденовирусы человека отно-
Adenovirus keratoconjunctivitis: causativeagent,epidemiology, clinical manifestations and treatment
A.N. Lukashev
Chumakov Institute of Poliomyelitis and Viral
Encephalitides
Abstract
Adenoviruses are well studied experimentally, but remain less explored from a clinical perspective. Adenoviruses rarely cause a serious disease, but are an important cause of relatively mild infections. In particular, adenoviruses are one of the key causes of conjunctivitis. The disease usually resolves spontaneously without complications, but lingers for 3–6 months, and the officially registered incidence does not reflect the social and economic impact of the disease. To date there is no specific therapy for adenovirus keratoconjunctivitis with a proven efficacy. This review covers molecular properties of adenoviruses, their clinical relevance, epidemiology, clinics and treatment of adenovirus keratoconjunctivitis.
Introduction
Adenoviruses are DNA viruses capable of infecting humanandothervertebrates.Adenoviruseswerefirst isolated in 1953 from adenoid (nasopharyngeal tonsil) tissue, hence the name. Studies on adenoviruses had a big impact on the development of molecular biology in general. mRNA splicing was first discovered in 1975 in adenoviruses. Adenoviruses have also been the first model to study viral oncogenesis. Relative ease of genome modification, culturing and purification led to wide use of adenovirus vectors for gene therapy, making them the second most popular after lentivirus vectors.
Classification of adenoviruses
Adenoviruses comprise the family Adenoviridae. To date there are over 100 known adenoviruses isolated from diverse vertebrates. Genus classification is based upon the host species due to strict host specificity of adenoviruses. Human adenoviruses belong to the genus Mastadenovirus

4 |
|
|
|
|
Медицинскаявирусология.томXXVII(2) |
|
сятся к роду Mastadenovirus (аденовирусы млеко- |
|
(mammalian adenoviruses) and are further |
||||
|
||||||
питающих) и подразделяются на 7 видов (Табл. 1). |
|
subdivided into 7 species (Table 1). Some species |
||||
В некоторые из видов аденовирусов человека так- |
|
of human adenoviruses also include simian |
||||
же входят аденовирусы, выделенные от приматов |
|
adenoviruses (not shown in the table). |
||||
(в таблице не приведены). |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Таблица 1. Классификация аденовирусов человека. |
|
|
|
|
|
|
Table 1. Classification of human adenoviruses. |
|
|
|
|
|
|
(http://www.ncbi.nlm.nih.gov/taxonomy). |
|
|
|
|
|
|
|
||
Вид |
Серотипы /Types |
Онкогенный потенциал. / |
Основные заболевания. / |
|
||
Oncogenic potential |
Major clinical manifestations |
|
||||
|
|
|
||||
A |
12, 18, 31, 61 |
Высокий /High |
|
|
||
|
|
|
|
|
|
|
B |
3, 7, 11, 14, 16, 21, 34, 35, 55 |
Умеренный / Moderate |
ОРВИ, пневмония / Acute |
|
||
respiratory infection, pneumonia |
|
|||||
|
|
|
|
|
|
|
C |
1, 2, 5, 6, 57 |
Низкий или отсутствует / |
ОРВИ /Acute upper respiratory |
|
||
Low or absent |
infection |
|
||||
|
|
|
||||
D |
8, 9, 10, 13, 15, 17, 19, 20, 22-30, 32, 33, |
низкий или отсутствует / |
керато-конъюнктивит / |
|
||
36-39, 42-49, 51, 53, 54, 58, 60, 62-67 |
Low or absent |
Keratoconjunctivitis |
|
|||
E |
4 |
Неизвестен / Not known |
Пневмония / Pneumonia |
|
||
|
|
|
|
|
||
F |
40, 41 |
Неизвестен / Not known |
Гастроэнтерит / Gastroenteritis |
|
||
|
|
|
|
|
||
G |
52 |
Неизвестен / Not known |
Гастроэнтерит / Gastroenteritis |
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Строение аденовирусов |
|
|
Structure of adenoviruses |
|||
Аденовирусы не имеют липидной оболочки, |
|
Adenoviruses do not have a lipid envelope, the |
||||
капсид икосаэдрической формы, размером око- |
|
capsid is icosahedric, sized ca. 90 nm (Fig. 1). The |
||||
ло 90 нм (Рис. 1). Капсид состоит из 240 копий |
|
capsid is comprised by 240 copies of hexon protein, |
||||
белка гексона (Hexon), 12 копий пентона (Pen- |
|
12 copies of penton protein at icosahedre apices |
||||
ton) в вершинах икосаэдра и 12 «нитей» – три- |
|
and 12 fibers (trimmers of the fiber protein) linked to |
||||
меров белка фибер (Fiber), связанных с пентона- |
|
pentons. Inside the capsid there is the matrix protein |
||||
ми. Внутри капсида находится матричный белок |
|
C, the viral genome and several accessory proteins |
||||
С, вирусный геном и ряд вспомогательных бел- |
|
(Berk, 2007). |
|
|
||
ков (Berk, 2007). |
|
|
|
|
|
|
Рис.1. Структура аденовирусного капсида
(Gupta and Hodge, 2006)
Fig.1. Structure of the adenovirus capsid (Gupta and Hodge, 2006)

medical virology. vol. XXVII (2) |
5 |
|
Ранние этапы инфекции |
|
Early stages of infection |
|
||
Проникновение вируса в клетку начинается со |
|
Virus entry into a cell begins with interaction |
связывания специфического рецептора в голов- |
|
of a specific receptor in the Fiber knob with the |
ке фибера с клеточным рецептором (у всех адено- |
|
cellular receptor (in all adenoviruses except for |
вирусов кроме вида В – коксаки-адено рецептор, |
|
species B – so-called coxsackie-adeno receptor, |
CAR). Затем происходит связываниевторого рецеп- |
|
CAR). Then the second receptor at the penton base, |
тора, расположенного в основании пентона, (мо- |
|
the RGD motif, binds to integrins at the cell surface |
тив RGD) с интегринами на поверхности клетки для |
|
to provide virus internalization into the endosome |
интернализации вируса в эндосому (Рис. 2). После |
|
(Fig. 2). After partial disassembly, the capsid exits |
частичной разборки капсида, он выходит из эндо- |
|
the endosome and is transported to the nuclear |
сомы и транспортируется к ядерной поре по си- |
|
pore by the microtubule system. Viral DNA enters |
стеме микротрубочек. Вирусная ДНК проникает в |
|
the nucleus via the nuclear pore, then early viral |
ядрочерезядернуюпору,затемтранскрипцияран- |
|
mRNAs are transcribed by the cellular transcription |
нихРНКосуществляетсясистемойклеточнойтранс- |
|
system (Berk, 2007). |
крипции (Berk, 2007). |
|
|
|
|
|
Рис. 2. Ранние этапы репликации аденовируса (Greber, 2002) Fig. 2. Early events of adenovirus replication cycle (Greber, 2002)
Реализация генома
Геном аденовирусов представлен линейной двухцепочечной ДНК размером около 35 тысяч пар оснований.На5’концахсгеномнойДНКсвязантерминальный белок (ТР). Геном достаточно сложно устроен, кодирует большой набор белков под контролем5ранних(E1A,E1B,E2,E3,E4),трехотложенных (IX, IVa2 и поздний E2) и одного позднего промотера (L), с которого транскрибируются 5 поздних РНК(L1-L5)(Рис.3)(Berk,2007).Вирусныебелкимо-
гут кодироваться на основной и комплементарной цепях ДНК. В результате сплайсинга аденовирусы могут производить варианты вирусных белков с общей и вариабельными частями. Лучше всего изучены белки вида Human adenovirus C потому, что аденовирус2типаявляется«прототипным»аденовирусомчеловека,ааденовирус5типабылосновойдля большей части аденовирусных векторов, используемыхвбиотехнологии.Сплайсингбелковдругихвидов аденовирусов изучен хуже, и экспериментальногодоказательстваналичиягомологоввсехбелков аденовируса 5 типа и их роли в репликации других аденовирусовврядеслучаевнет.
Genome strategy
Adenovirus genome is a linear double-stranded DNA of about 35 kilobases. The Terminal protein (TP) is bound to the 5’ termini of the genomic DNA. The genome is rather complex and encodes for a large set of viral proteins under control of 5 early (E1A, E1B, E2, E3, E4), three delayed early (IX, IVa2 and late E2) and one major late promoter that controls transcription of five late mRNAs (L1L5) (Fig. 3) (Berk, 2007). Viral proteins can be encoded on the main and on the complementary genome strand. After splicing, adenoviruses can produce variants of viral proteins with common and variable parts. Proteins of the species Human adenovirus C are the best studied because Human adenovirus 2 (HAdV-C2) is the prototype human adenovirus, and HAdV-C5 was the basis of most of the adenoviral vectors used in biotechnology. Splicing of proteins of other adenovirus species is less studied, and there is no experimental proof that homologs of all HAdV-C5 are produced by other adenoviruses.
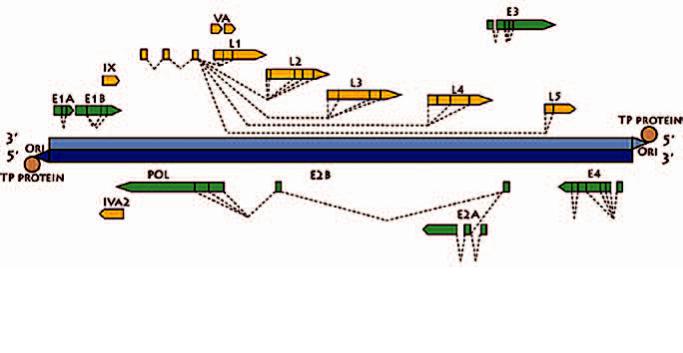
6 |
Медицинскаявирусология.томXXVII(2) |
Рис.3.Общая схема реализации аденовирусного генома
Fig.3. General overview of the adenovirus genome strategy (http://viralzone.expasy.org)
Белки, кодируемые под контролем позднего |
|
Proteins encoded under the major late |
|
||
промотера, имеют гомологи у всех аденовирусов. |
|
promoter have homologs in all adenoviruses. |
Многие ранние белки обнаруживаются только у |
|
Many early proteins are found only in the |
аденовирусов млекопитающих. Наибольшей вари- |
|
mammalian adenoviruses. Proteins expressed |
абельностью между видами аденовирусов отлича- |
|
under E3 promoter are the most variable between |
ются белки под контролем Е3 промотера. |
|
adenovirus species. |
Белки аденовирусов |
|
Adenovirus proteins |
Два белка, экспрессируемых под контролем |
|
Two proteins expressed under the E1A |
промотера Е1А, отвечают за переход клетки в S |
|
promoter control transition of the cell into the S |
фазу клеточного цикла и активацию транскрипции |
|
phase of the cell cycle and activation of viral mRNA |
вирусных мРНК. Два белка Е1В блокируют апоптоз. |
|
transcription. Two E2 proteins block apoptosis. E2 |
Под контролем промотера Е2 кодируется вирус- |
|
promoter region encodes for the viral polymerase |
ная полимераза и белки репликативного комплек- |
|
and replication complex proteins. E3 proteins |
са.БелкиЕ3отвечаютзаблокированиеинтерферо- |
|
block interferon response and suppress innate |
новогоответаиподавлениеврожденногопротиво- |
|
antiviral immunity of the host. E4 block encodes |
вирусного иммунитета хозяина. В блоке Е4 кодиро- |
|
for accessory proteins that have various functions: |
ваны вспомогательные белки, выполняющие раз- |
|
enhancement of transcription and translation, |
ные задачи – активация транскрипции и трансля- |
|
suppression of apoptosis etc. Late proteins are |
ции, подавление апоптоза и т.д. Поздние белки яв- |
|
structural proteins, take part in virus assembly and |
ляются структурными белками, участвуют в сборке |
|
initiate cell death to release mature virions (Berk, |
вируса и запускают гибель клетки для обеспечения |
|
2007). |
выхода зрелых вирионов (Berk, 2007). |
|
|
Вариабельность генома аденовирусов |
|
Variability of adenovirus genome |
Аденовирусный геном представлен двухцепо- |
|
Adenovirus genome is a double stranded DNA, |
чечной ДНК, а репликативный комплекс вируса |
|
and the replicative complex of the virus has error |
имеет функцию коррекции ошибок. Эти факторы |
|
correction capacity. These factors result in relatively |
обуславливают достаточно высокую стабильность |
|
high stability of the adenovirus genome. Substitution |
аденовирусного генома. Частота накопления мута- |
|
rateinadenovirusesisabout5*10-5 substitutions/site/ |
ций у аденовирусов составляет около 5*10-5 замен |
|
year, i.e. 100 times lower than in most of RNA and |
на сайт в год, т.е. примерно в 100 раз ниже, чем у |
|
small DNA viruses (Firth et al., 2010). Adenoviruses |
большинства РНК-содержащих вирусов и мелких |
|
feature both point mutations and indels, especially |
ДНК-содержащих вирусов (Firth et al., 2010). У аде- |
|
in hypervariable parts of the genome. The apparent |
новирусовнаблюдаютсякакточечныемутации,так |
|
genetic variability among natural isolates largely |
и вставки/делеции, особенно в гипервариабель- |
|
depends upon prevalence of that species and type, |

medical virology. vol. XXVII (2) |
7 |
ных участках генома. Наблюдаемая генетическая вариабельность природных изолятов аденовирусов в значительной степени зависит от распространенности данного вида и серотипа и от числа геномов, представленных в базе данных GenBank, т.е. вариабельность редких аденовирусов может быть значительно недооценена. Различные участки аденовирусного генома характеризуются разной вариабельностью. Наибольшее разнообразие нуклеотидной последовательности наблюдается в участках генома, кодирующих поверхностные белки, в особенности их участки, образующие серологические эпитопы на поверхности вируса (Рис. 4). В участках генома, кодирующих фибер и гексон, вариабельность нуклеотидной последовательности между серотипами одного вида может достигать
50% (Lukashev et al., 2008). Участки генома, коди-
рующие неструктурные белки, значительно более консервативны (Рис. 4). Вариабельность аминокислотной последовательности в пределах вида может практически отсутствовать, однако синонимические замены встречаются, и вариабельность нуклеотиднойпоследовательностисоставляетпоряд-
ка 3-5% (Lukashev et al., 2008).
and upon the number of genomes represented in Genbank, i.e. variability of rare adenoviruses may be largely underestimated. Distinct regions of the adenovirus genome feature different variability. The highest variability is observed in regions that encode forthesurfaceproteins,especiallyatthelocationsthat correspond to the serological surface epitopes (Fig. 4). Genome regions that encode for fiber and hexon can vary by up to 50% nucleotide sequence between distinct serotypes of the same species (Lukashev et al., 2008). Genome regions that encode for nonstructural proteins are much more conserved (Fig. 4). Variability of the amino acid sequence may be almost absent, however synonymous substitutions occur and the nucleotide sequence variability may be around 3-5% (Lukashev et al., 2008).
Рис.4. Средние нуклеотидные дистанции между геномами аденовирусов видов C и D. Модель замен Кимура, окно 500 нт, шаг окна 20 нт (Lukashev et al., 2008)
Fig.4. Mean nucleotide distances between genomes of Human adenovirus C and D species. Kimura substitution model, window 500 nt, step 20 nt (Lukashev et al., 2008)
8 |
Медицинскаявирусология.томXXVII(2) |
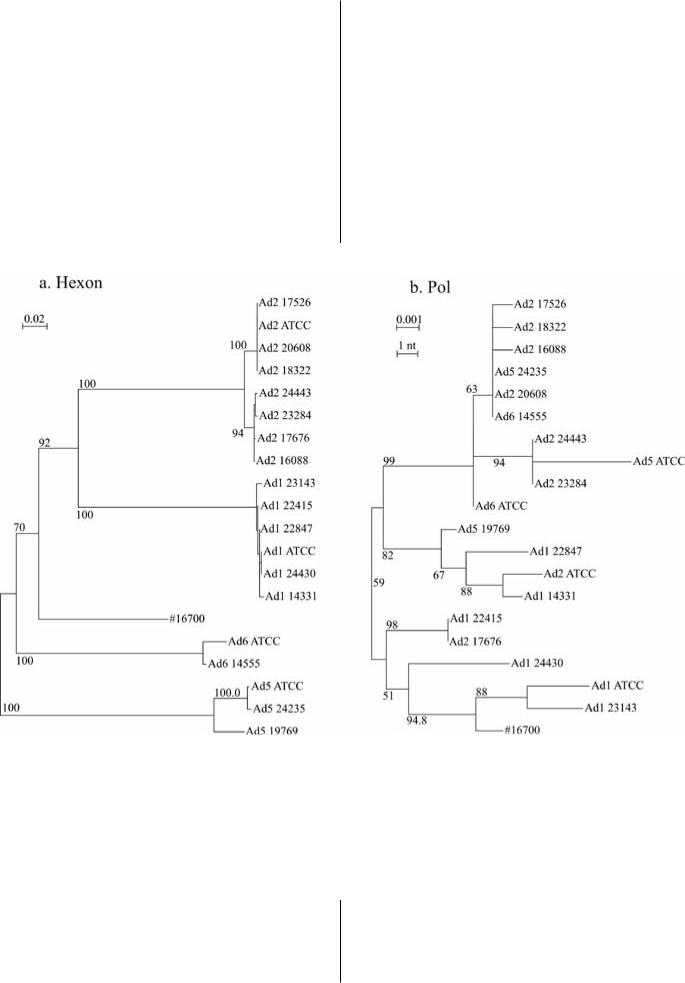
Кроме мутаций и вставок/делеций, важным механизмом эволюции аденовирусов является рекомбинация. Возможность рекомбинации у аденовирусов была открыта более 40 лет назад (Williams and Ustacelebi, 1971). Долгое время рекомбинация считалась лабораторным феноменом. В 2008 году было показано, что рекомбинация часто происходит в неструктурной области генома между аденовирусамиодноговида(Lukashevet al.,2008).Филогенетическое группирование соответствует серотипу только в областях генома, кодирующих поверхностныебелкикапсида,авдругихобластяхгенома аденовирусы разных типов «перемешаны» в пределах вида (Рис. 5)
In addition to point mutations and indels, recombination is an important mechanism of adenovirus genome variability. Possibility of recombination in adenoviruses was discovered over 40 years ago (Williams and Ustacelebi, 1971). For a long time recombination was perceived as a laboratory phenomenon. In 2008 it was shown that recombination commonly occurs in the non-structural genome regions between adenoviruses of the same species (Lukashev et al., 2008). Phylogenetic grouping corresponds to the serotype only in the genome regions that encode for the surface proteins of the capsid, while in other genome regions adenoviruses of different types are shuffles within a species (Fig. 5)
Рис.5. Филогенетическое группирование аденовирусов вида С в двух областях генома, гексоне (позиции генома 19198-19930) и полимеразе (позиции генома 7340–8466) (Lukashev et al., 2008). Деревья построены по алгоритму Neighbor-joining с моделью замен Kimura. Цифры в узлах дерева – процент bootstrap-псевдорепликатов, поддерживающих данное группирование
Fig.5. Phylogenetic grouping of adenovirus C in two genome regions, Hexon (nt 19198–19930) and polymerase (nt 7340–8466) (Lukashev et al., 2008). Neighbor-joining trees, Kimura substitution model. Numbers at tree nodes are percent of bootstrap pseudo-replicates that supported that grouping
В результате геномы аденовирусов являются сложномозаичными рекомбинантами относительно других вирусов вида (Рис. 6), и в областях генома, кодирующих неструктурные белки, нуклеотидная последовательность аденовируса определен-
As a result, adenovirus genomes are complex mosaic recombinants relative to other viruses of the species (Fig. 6), and in the non-structural genome regions nucleotide sequence of an adenovirus may correspond to any possible nucleotide sequence
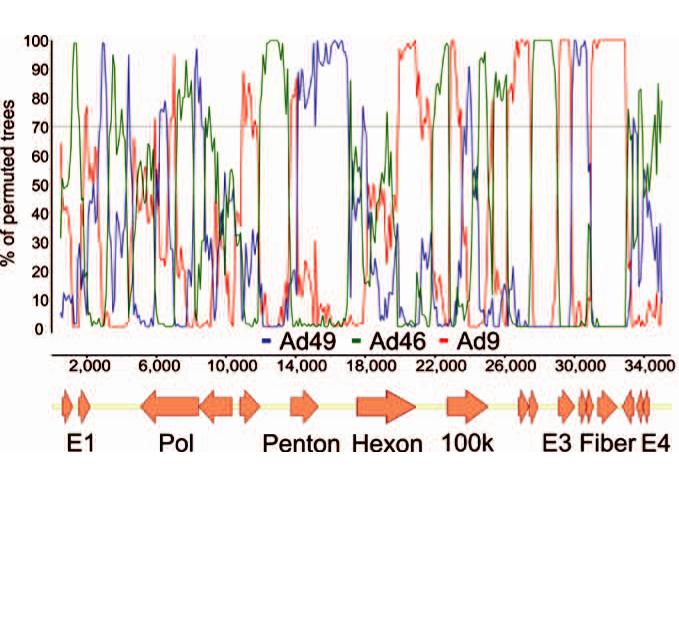
medical virology. vol. XXVII (2) |
9 |
|
ного серотипа может соответствовать любой воз- |
|
within a species (Lukashev et al., 2008). Such chimeric |
|
||
можной нуклеотидной последовательности в пре- |
|
viruses can as well cause keratoconjunctivitis (Aoki et |
делах всего вида (Lukashev et al., 2008). Такие хи- |
|
al., 2008). |
мерные аденовирусы могут быть и возбудителями |
|
|
кератоконъюнктивита (Aoki et al., 2008). |
|
|
|
|
|
Рис.6. Bootscan-анализ аденовируса 17 типа (представитель вида D, включающего возбудителей аденовирусного кератоконъюнктивита). Ось X – позиция в геноме, ось Y – процент bootstrap-псевдорепликатов, поддерживающих филогенетическое группирование аденовируса 17 типа с вирусами сравнения, указанными под графиком
(Lukashev et al., 2008)
Fig.6. Bootscan-analysis of adenovirus type 17 (representative of Adenovirus D species that includes causative agents of keratoconjunctivitis) X axis – genome position, Y axis – percent of bootstrap pseudo-replicates that support grouping of HAdV-D17 with query adenoviruses indicated below the graph (Lukashev et al., 2008)
Патогенность аденовирусов для человека |
|
Pathogenicity of adenoviruses for humans |
|
||
Аденовирусы достаточно часто обнаруживаются |
|
Adenoviruses are frequently found in upper |
в верхних дыхательных путях у здоровых лиц. У де- |
|
respiratory tract of healthy humans. In children |
тей аденовирусы определяются в гиперплазирован- |
|
adenoviruses are found in 47% of adenoid samples |
ных аденоидах в 47% случаев (Proenca-Modena et |
|
(Proenca-Modena et al., 2012). About 1% of |
al., 2012). Около 1% здоровых детей выделяют аде- |
|
healthy children excrete adenoviruses with feces. In |
новирусы с фекалиями, причем в организованных |
|
organized groups and in immunodeficient children |
коллективах и у детей с иммунодефицитами часто- |
|
asymptomaticexcretionratescanreach50%(Ivanova |
табессимптомноговыделенияаденовирусовможет |
|
et al., 2012). Due to high asymptomatic prevalence it |
достигать50%(Ivanovaet al.,2012).Поэтойпричине |
|
is difficult to precisely establish the etiological role |
оценить точную частоту заболеваний верхних дыха- |
|
of adenoviruses in gastroenteritis and respiratory |
тельныхпутейигастроэнтеритов,вызванныхадено- |
|
tract infections. Adenoviruses can also affect lower |
вирусами, затруднительно. Кроме того, аденовиру- |
|
respiratory tract and cause pneumonia. It was |
сымогутвызыватьпоражениенижнихдыхательных |
|
estimated that adenoviruses cause (or are involved |
путей(пневмонию).Считается,что аденовирусы вы- |
|
into combined infection cases) 5-10% of acute lower |
зывают (или вовлечены в полиэтиологические слу- |
|
respiratory infection cases in children (Brandt et al., |