

6.4. Изменения мышечной ткани в процессе старения
С увеличением возраста в мышечных клетках уменьшается содержание калия и растет концентрация натрия и хлора в связи со снижением активности ионных насосов. Ухудшается способность саркоплазматического ретикулума к связыванию и освобождению кальция.
Число рецепторов гладкомышечных клеток ко многим гормонам и медиаторам падает. Но при этом сродство, а следовательно, чувствительность ряда рецепторов к гормонам повышается.
Волокна поперечнополосатых мышц истончаются и уменьшаются в объеме, в них снижается число пучков миофибрилл, менее четко выявляется поперечная исчерченность. Увеличивается доля коллагеновых волокон б межмышечной соединительной ткани, что является причиной снижения эластичности мышц. Падают возбудимость миоцитов и амплитуда их потенциалов действия.
Однако при регулярных двигательных нагрузках активируется синтез многих белков, а размеры клеток не только скелетных, но и сердечной мышц, а также их митохондрий увеличиваются. Это приводит к улучшению сократительной активности миокарда, наполнения камер сердца и улучшению кровоснабжения тканей. Одновременно такие нагрузки приводят к поддержанию умственной работоспособности, преобладанию положительных эмоций. Эти механизмы лежат в основе профилактического действия регулярных физических нагрузок на организм пожилых и старых людей. Наблюдения в течение многих лет над 320 000 мужчин семи стран (США, Японии, Финляндии и др.) показало, что физические упражнения в часы досуга на V5 уменьшают риск развития болезней сердца и кровеносных сосудов.
V Глава 7 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
7Л. Функции центральной нервной системы
Центральная нервная система (ЦНС) в организме выполняет интегрирующую роль. Она объединяет в единое целое все ткани, органы, координируя их специфическую активность в составе целостных гомеостатических и поведенческих функциональных систем (П.К.Анохин). Основными частными функциями ЦНС являются следующие.
1. Управление деятельностью опорно-двигательного аппарата. ЦНС регулирует тонус мышц и посредством его перераспределения поддерживает естественную позу, а при нарушении восстанавливает ее, инициирует все виды двигательной активности (физическая работа, физкультура, спорт, любое перемещение организма).
2. Регуляция работы внутренних органов
осуществляется посредством вегетативной нервной системы и эндокринными железами:
а) в покое —- обеспечение гомеостазиса (по стоянства внутренней среды организма);
б) во время работы — приспособительная ре гуляция деятельности внутренних органов согласно потребностям организма и поддер жание гомеостазиса. Интенсивность функцио нирования отдельных органов изменяется в широком диапазоне и соответствует потреб ностям организма в данный момент. Функ ция одних органов может угнетаться, других, напротив, — стимулироваться. Например, при увеличении физической нагрузки дея тельность сердечно-сосудистой и дыха тельной систем усиливается, а желудочно- кишечного тракта — тормозится. С помощью вегетативной нервной системы регуляторные
94
волокнах типа С связана с особенностями проведения возбуждения в безмиелиновых волокнах. Лабильность также уменьшается от волокон Аа до С и находится в обратной зависимости от продолжительности фазы абсолютной рефрактерности. Возбудимость тоже уменьшается от волокон Аа (наибольшая возбудимость) к волокнам С (наименьшая возбудимость). Например, пороговая сила электрического тока у волокон С в 30—50 раз больше, чем у волокон Аа. Исследование факторов, блокирующих нервную проводимость, показало, что к давлению наиболее чувствительны волокна А, к кислородному голоданию (гипоксии) — волокна В, к местным анестетикам — волокна С.
Нервные волокна имеют две основные функции — проведение возбуждения и транспорт веществ, обеспечивающих трофическую функцию.
5.2.2. МЕХАНИЗМ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВНОМУ ВОЛОКНУ
Биопотенциалы могут быть локальными (местными), распространяющимися с декрементом (затуханием) на расстояние, не превышающее 1—2 мм, и импульсными (ПД), распространяющимися без декремента по всей длине волокна — на несколько десятков сантиметров, например от мотонейронов спинного мозга по всей длине нервного волокна до мышечных волокон конечностей с учетом длины самой конечности.
А. Распространение локальных потенциалов. Дакальные_потенциалы (препотенциал, рецепторный потенциал^ возбуждающий постсинаптический потенциал — ВПСП) изг меняют мембрандый_цо^ендиал--йокоя^_как правило^"в'сторону д&полярнзадии_в_2еэ^А~ тате входа в клетку Na+ согласно электрохимическому градиенту. В результате этого между участком волокна^ в .котором возник лотсштьтпттгбтенциал, и соседними участка-1 ми мембраны формируется электрохимический градиент, вызывающий передвижение, ионов. В частности, вошедшие в клетку ионы Na+ Начинают перемещаться в соседние участки, а ионы Na+ на наружной поверхности клетки движутся в противоположном наг правлении. В итоге поляризация мембраны соседнего участка уменьшится. Фактически это означает, что локальный потенциал из первичного очага распространился на соседний участок мембраны. Он затухает на расстоянии 1—2 мм от очага первичной деполяризации, что связано с отсутствием ионных управляемых каналов на данном участке
мембраны или неактивацией управляемых ионных каналов, продольным сопротивлением цитоплазмы волокна и шунтированием тока во внеклеточную среду через каналы утечки мембраны.
_Если возникшая деполяризация мембраны не сопровождается изменением проницае- мости потенциалзависимых натриевbiXj каль циевых и калиевых каналов, такую деполяри зацию называют, электротонической. Элек- '1 ротон ичее кое распросч ранение возбужде ния — физический механизм, оно характерно для тех фрагментов мембран возбудимых кле ток, где нет потенциалзависимых ионных ка налов. Такими участками являются, напри мер, большая часть мембраны дендритов нервных клеток, межперехватные промежут ки в миелиновых нервных волокнах. Если м_естный__ДО1енциал (рецепторный или Т5ПСП), распространяясь электратонически^ достигает участков__мембраны, способных ге нерировать ПД (аксонный холмик, перехва ты Ран вье, часть мембраны дендритов и?_воз- ' можно, сомы), но его амплитуда'прйГэтом не достигнет ЩШГ
критического ци^'тотакоЙ.потенциал называют претютен-циадо-м^-В его возникновении и распространении частично участвуют потенциалзависи-мые ионные каналы, однако при этом нет регенеративной (самоусиливающейся) деполяризации, характерной для ПД. Поэтому распространение такого потенциала происходит с затуханием амплитуды. Если легальный потенциал достигает участков мембраны, способных генерировать ПД, и его амплитуда выходит на критический уровень деполяризации, формируется ПД, который распространяется по всей длине нервного волокна без затухания.
Эффективность электротонического распространения биопотенциалов зависит от физических свойств нервного волокна — сопротивления и емкости его мембраны, сопротивления" цитоплазмы. Электротолинес-кое- проведение в нервном волокне улучшается^ при^увеличении его диаметра, что связа-но^с-уменьшением сопротивления^щщ^пЯаз;: мы, а также при миелинизации волокна, увеличивающей сопротивление
2) й уменьшающей ее" емкость (до_ 0,005 мкФ/см2). ЭффeJaщн^cть_эл5ктJютo-нического проведен_ия_хаЁактеризует постоянная длины мембраны (Хт). Это;_"р_асстоя--ние,. на которое может электротонически распространиться бйопотенЩШ;—fmKa~-&ro амплитуда-неуменьшится до 37~"% ОТ UCxuu^j-ной^вели^ины, Постоянная длины для локальных потенциалов реально не превышает
68
А — возникновение ПД в нервном волокне; Б — распространение ПД в обе стороны от места возникновения. 1 — область деполяризации; 2 — соседние области, 3 — область реполяризации на месте предшествующего ПД.

Для передачи возбуждения^ на большие расстояния н^^хошшх)~^оринрошаия£,-ПЛ^ В его""расп"ространении, кроме электротонического механизма, существенная рольлфи-надлеэкит механизму регенеративной деполяризации, позволяющей сохранить амплитуду ПД на всем пути его следования.
Б. Проведение потенциала действия осуществляется с использованием как физического (электротонического), так и физиологического механизмов. Обязательным условием проведения нервного импульса является наличие на всем протяжении или в ограниченных, но повторяющихся участках волокна потенциалзависимых ионных каналов^-ситйет-ственных за формирование ПД.„ В распространении ПД можно выделить Два..ДГДДа-этап электротонического проведения, обусловленный физическими свойствами нервного волокна, и эдж^шеаЩииЛД-Р новом участке на пути его д в иже ншГ, "обусловлен -ный реакцией ионных каналов. В зависимости от расположения и концентрации ионных каналов в мембране волокна возможно два типа проведения ПД: непрерывный и сальта-торный (скачкообразный).
 11Д
осу-
11Д
осу-
волокнахдш&а. распределение по- й^н~нь1^канадав .^^acj^yj: ^^оведение нервного импульса начинается с этапа электротонического распространения возникшего ПД. Амплитуда ПД нервного волокна (мембранный потенциал + инверсия) составляет око 90 мВ, постоянная длины мембраны (^) безмиели новых волокнах] равна 0,1 — 1,0 мКГ
Поэтому ПД, распространяясь на этом расстоянии ка^^^кт£От0ни^сктай"П оте н ци ал^и "сохранив как минимум 37 % своей амллиту-ДБгрспосббён дeпoляpизoвaть_Jrfgм_бpaнy до кртггтГчесКОто уровня и генерировать на всем протяжении новые ПД,(рис. 5.3). При этом на этапе электротонического распространения нервного импульса ионы движутся вдоль волокна между деполяризованным и поляризованным участками, обеспечивая проведение возбуждения в соседние участки волокна. Реально при неповрежденном нервном волокне этап чисто электротонического распространения ПД (вдоль мембраны) предельно мал, так как потенциалзависимые каналы имеются в непосредственной близости друг от друга и, естественно, — от возникшего потенциала действия и наблюдается только до достижения деполяризации, равной 50 % Екр.. Далее включается перемещение ионов в клетку (нервное волокно) и из клетки за счет активации ионных каналов.
При формировании нового ПД в соседнем участке в фазе деполяризации возникает мощный ток ионов натрия в клетку вследствие активации натриевых каналов, приводящий к регенеративной (самоусиливающейся) деполяризации. Этот ток обеспечивает формирование нового ПД той же амплитуды, представляющий собой, как обычно, сумму двух величин — мембранного потенциала покоя и инверсии. В связи с этим проведение ПД осуществляется без декремента (без снижения амплитуды). Таким образом, непрерывное распространение нервного импульса идет через генерацию новых ПД по эстафете, когда каждый участок мембраны выступает сначала как раздражаемый (при поступлении к нему электротонического потенциала), а
 69
69

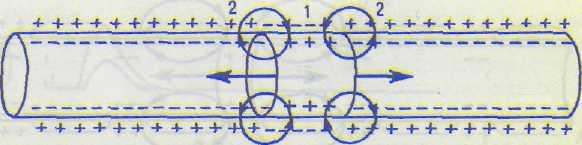
А — возникновение ПД в узловом перехвате среднего участка волокна; Б — распространение ПД в обе стороны от места его возникновения; стрелками показаны токи на электротоническом этапе распространения ПД.
затем как раздражающий (после формирования в нем нового ПД).
2![]() .
Сащпшпорный
тип проведения нер_вно-го
импульса осущестъляется^ в миелиновых
волокнахТтипьГАи
В^ ^£ терна
_ ко^щештдшт^ ионных
каналов только ^ ках__мембраяыЦв1п|рехватах
Еанвье)! где их плотность
достигает 12 000 на 1 мкм2,
что примерно
в 100 раз выше, чем в мембранах безмиелиновых
волокон. Bj^Gj^cjjiJsiJiejutiiOj:
вых
муфт (межуздовых сегменте в К_обладдЮ-щих
хорошими иппл^ру|ошими_свойствами.
потенциалзависимых
каналов, почти.. HeTj
и мембрана
осевого цилиндра там практически
невозбудима.
В этих условиях ПД,
возник-ший
в одном перехвате Ранвье, эдектротони-чёски
(вдоль волокна, без участия ионных
каналов)
распространяется до соседнего перехвата,
деполяризуя там мембрану до критического
УРОВНЯ, ЧТО ГДтиОЛИТ К ВОЗЦ.ИКНО-
.
Сащпшпорный
тип проведения нер_вно-го
импульса осущестъляется^ в миелиновых
волокнахТтипьГАи
В^ ^£ терна
_ ко^щештдшт^ ионных
каналов только ^ ках__мембраяыЦв1п|рехватах
Еанвье)! где их плотность
достигает 12 000 на 1 мкм2,
что примерно
в 100 раз выше, чем в мембранах безмиелиновых
волокон. Bj^Gj^cjjiJsiJiejutiiOj:
вых
муфт (межуздовых сегменте в К_обладдЮ-щих
хорошими иппл^ру|ошими_свойствами.
потенциалзависимых
каналов, почти.. HeTj
и мембрана
осевого цилиндра там практически
невозбудима.
В этих условиях ПД,
возник-ший
в одном перехвате Ранвье, эдектротони-чёски
(вдоль волокна, без участия ионных
каналов)
распространяется до соседнего перехвата,
деполяризуя там мембрану до критического
УРОВНЯ, ЧТО ГДтиОЛИТ К ВОЗЦ.ИКНО-
вению нового ПД, т.е. возбуждение проводится скачкообразно (рис. 5.4). Постоянная длина мембраны миелинового волокна достигает 5 мм. Это значит, что ПД, распространяясь электротонически на этом расстоянии, сохраняет 37 % своей амплитуды (около 30 мБ) и может деполяризовать мембрану до критического уровня (пороговый потенциал в перехватах Ранвье равен около 15 мВ). Поэтому в случае повреждения ближайших на пути следования перехватов Ранвье потенциал действия может электротонически возбудить 2—4-й и даже 5-й перехваты. Сальтаторное проведение ПД по миелино-вым волокнам является эволюционно более поздним механизмом, возникшим впервые у позвоночных. Оно имеет два важных преимущества по сравнению с непрерывным прове-
дением возбуждения. Во-первых, оно более экономично в 3Hepj^ejjjta&uuQM^iiiaHЈ. т.к. воз-"о^ШГаются только перехваты Ранвье, площадь которых менее 1 % мембраны, и, следовательно, надо меньше энергии для восстановления трансмембранных градиентов Na+ и К+, уменьшающихся в процессе формирования ПД. Во-вторых, возбуждение .проводитсд_с большей скоростью (см. табл. 5.2), чем в без-"миелиновых волокнах, так как возникший ПД на протяжении миелиновых муфт распространяется электротонически, что (Д^0^раз_ёы-стрее^чем скорость непрерывного проведения ПД в безмиелиновом волокне.
5.2.3. ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ В НЕРВНЫХ СТВОЛАХ
В периферической нервной системе волокна объединены с помощью соединительнотканных оболочек в нервные стволы (нервы). В одном нерве могут быть тысячи нервных волокон: например, в срединном и мышеч-но-кожном нервах имеется 27—37 тыс. нервных волокон. Волокна в нервах могут быть миелиновыми и безмиелиновыми, афферентными и эфферентными. В естественных условиях каждое волокно нерва возбуждается от своего источника (например, эфферентное — от аксонного холмика, афферентное — от рецептора), и ПД в них проводятся асинхронно. Кроме того, чувствительные и двигательные волокна проводят импульсы в противоположных направлениях. Суммарная электрическая активность нерва создается электрической активностью составляющих его волокон и зависит от числа возбужденных волокон, степени шунтирования местных токов невозбужденными волокнами,
70
 А ВС
А ВС
Типы волокон

о
Рис. 5.5. Биоэлектрическая активность раздражаемого нерва при монополярном (I, П) и биполярном (III) отведении (по Д.П.Матюшкину, 1984, с изменениями).
РЭ — раздражающие электроды; ОЭ — отводящие электроды и регистрирующее устройство; П — участок повреждения; Г — суммарный потенциал нерва при малом расстоянии проведения: два потенциала на действие раздражителей возрастающей силы (закон силы); II — дисперсия суммарного потенциала при большом расстоянии проведения; Ш — формирование двухфазного суммарного потенциала из однофазных потенциалов. Mi, M2 — монополярные отведения; Б — биполярное отведение-
СИНХрОННОСТИ проведения ПД в волокнах. В связи с этим анализ суммарной электрической активности нерва (нейрограммы) представляет трудную задачу.
В лабораторных условиях при монополярном отведении, когда один электрод расположен на неповрежденном участке нерва, а второй — на поврежденном (деполяризованном) участке, можно зафиксировать суммарный монофазный ПД нерва и его дисперсию (расслоение) во времени. Если отводящий электрод расположен близко (до 3 мм) к раздражающему, через который подают сильный одиночный стимул (рис. 5.5, М,), то регистрируется суммарный ПД нерва, напоминающий по форме ПД отдельного нервного волокна, но растянутый по времени (см. рис. 5.5, I). Суммарный ПД нерва в отличие от ПД отдельного волокна не подчиняется закону «все или ничего». Это означает, что при увеличении силы раздражения увеличивается число возбужденных нервных волокон: в возбуждение вовлекаются, кроме Аа-волокон, менее возбудимые Ар-, Ay-, A6-, В-волокна и, наконец, наименее возбудимые С-волокна (закон силовых отношений — увеличение ответной реакции с увеличением силы раздражения).
Если отводящий электрод расположен на достаточном удалении от раздражающего электрода (до 80—100 мм, см. рис. 5.5, М2), то фиксируется расслоение суммарного ПД нерва на несколько пиков соответственно типам нервных волокон (см. рис. 5,5,11). Это связано с неодинаковой скоростью проведения ПД в разных волокнах нерва: сначала до места регистрации доходят нервные импульсы по быстропроводящим Аа-волокнам, через некоторое время по Ар, затем по Ау и т.д. Позже всего до места регистрации доходят ПД по С-волокнам.
Если отведение биполярное и оба отводящих электрода расположены на неповрежденных участках нерва и недалеко от раздражающего электрода (чтобы избежать дисперсии потенциала, см. рис. 5.5, Б), то при сильном одиночном стимуле фиксируется двухфазный суммарный потенциал (рис. 5.5, III). Возникновение этих фаз связано с тем, что, когда волна возбуждения находится под первым (ближайшим к месту раздражения) отводящим электродом, этот участок становится электроотрицательным по отношению к покоящемуся участку под вторым отводящим электродом и луч осциллографа отклоняется вверх. Когда же волна возбуждения доходит
71
до второго электрода, а под первым электродом мембраны волокон уже реполяризованы, то луч осциллографа отклоняется в противоположную сторону — вниз.
5.2.4. ХАРАКТЕРИСТИКА ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВНЫМ ВОЛОКНАМ