

Биотехнология в животноводстве
.pdf1.2.Реализация наследственной информации. Наследственная ин-
формация, закодированная в молекуле ДНК, реализуется на всех этапах жизнедеятельности клетки и многоклеточного организма в процессе био- синтеза. Исследования показали, что каждый ген контролирует синтез од- ного соответствующего фермента («один ген – один фермент») и реализа- ция наследственной информации осуществляется в процессе синтеза. Ген, локализованный на определенном участке молекулы ДНК, контролирует синтез первичной молекулы белка, представляющей собой полипептид- ную цепь, специфичность которой зависит от порядка чередования в ней аминокислот.
Белкам принадлежит исключительно важная роль в жизнедеятельно- сти каждой клетки и всего многоклеточного организма. Они участвуют в построении мембран, хроматина, рибосом, митохондрий, являются со- ставной частью сложных белков. В качестве ферментов и гормонов они управляют всеми процессами в клетке и в многоклеточном организме. Подавляющее большинство метаболических реакций, от которых зависит развитие признака или свойства, находится под контролем ферментов и, следовательно, генов.
Первичная молекула белка представляет собой цепочку, состоящую из 100-300 различных аминокислот и более, порядок чередования которых определяет специфичность данной молекулы: каждая из 20 аминокислот может встречаться многократно, но местонахожде-
ние контролируется ДНК. В настоящее время для многих молекул белка установлена их первичная структура, т.е. порядок чередования аминокис- лот в полипептидной цепи.
Вторичная структура белковой молекулы зависит от первичной: аминокислоты в полипептидной цепи соединяются водородными связями между NН – и СО – группами, в результате чего она свертывается в так называемую альфа-спираль. Образование больших альфа-
спиральных участков характерно для фибриллярных белков. В молекулах ферментов спиралеобразных участков значительно меньше.
Третичная структура белковых молекул образуется в результате связывания так называемыми дисульфидными мостиками (S-S) двух цистеиновых остатков аминокислот. Это определяет специфическое пространственное расположениеполипептидных цепей.
Четвертичная структурабелковых молекул характеризуется тем, что они состоят из двух-четырех различных, стабильно соединенных полипептидных цепей. Такая структура характерна для глобулярных
11
белков, в т.ч. для многих ферментов. Вторичная, третичная и четвертичная
структуры белковых молекул зависят от числа и порядка чередования аминокислот в полипептидной цепи, т.е. от первичной структуры.
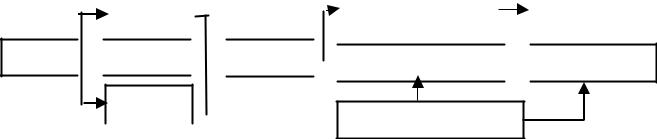
Процесс синтеза белка в клетке называется биосинтезом. Он осуществляется под контролем молекулы ДНК, которая таким образом реализует закодированную в ней наследственную информацию. Схема-
тично реализацию наследственной информации можно представить сле- дующимобразом:
и РНК |
|
Ферментный |
|
Свойство |
ДНК  т РНК
т РНК 
 Белок
Белок  Структурный
Структурный  Признак
Признак
р РНК 
 Предшественник
Предшественник
Крик в 1960 г. предложил матричную теорию синтеза белков, осно- ванную на трех положениях:
а) комплементарности азотистых оснований нуклеиновых кислот;
б) линейной последовательности их расположения в молекуле таких кислот;
в) и так называемом центральном постулате, заключающемся в том, что передача информации может происходить только с нуклеиновой кислоты на нуклеиновую или на белок; с белка же на белок передача невозможна.
Следовательно, матрицами для синтеза белка могут быть только нук- леиновые кислоты. Синтез ферментов и других белков, необходимых для развития и жизни организма, происходит в основном в первой половине интерфазы – стадии G1, т.е. до начала репликации ДНК. Для его осущест- вления необходимы: ДНК как матрица, на которой синтезируется ком- плементарная ей молекула м-РНК; м-РНК; рибосомы, состоящие из двух субчастиц, включающих рибосомную РНК и рибосомальные белки; набор т-РНК для каждой из 20 аминокислот, входящих в состав молекулы белка; аминокислоты и ферменты – РНК – полимераза, ферменты, регулирую-
щие начало и конец синтеза белковой молекулы и соединения в одну цепь аминокислотных остатков, а также ферменты, активизирующие амино- кислоты.
Процесс биосинтеза сложный и включает ряд этапов – транс-
крипцию, сплайсинг и трансляцию. При транскрипции последователь-
12

ность оснований в наружной цепи ДНК как бы «записывается» компле- ментарными им основаниями в синтезируемой на соответствующем уча- стке ДНК молекуле м-РНК и осуществляется в ядре, иначе говоря, транс-
крипция представляет собой запись в м-РНК последовательности оснований в ДНК.
Этот синтез осуществляется при участии комплекса ферментов, глав- ным из которых является ДНК – зависимая РНК –полимераза, которая прикрепляется к начальной (инициальной) точке молекулы ДНК, распле- тает двойную спираль и, перемещаясь вдоль одной из нитей, синтезирует рядом с ней комплементарную нить м-РНК. В результате транскрипции
мРНК содержит генетическую информацию в виде последовательного чередования нуклеотидов, порядок которых точно скопирован с соответ- ствующего участка (гена) молекулы ДНК.
Дальнейшие исследования показали, что в процессе транскрипции синтезируется так называемая про-мРНК- предшественник зрелой м- РНК, участвующей в трансляции. Про-мРНК имеет значительно большие размеры и содержит фрагменты, не кодирующие синтез соответствующей полипептидной цепи. В ДНК наряду с участками, кодирующими р-РНК, т-РНК и полипептиды, имеются фрагменты, не содержащие генетической информации. Они получили названия интронов в отличие от кодирующих фрагментов, которые называются экзонами.
Интроны обнаружены на многих участках молекул ДНК. Так, напри- мер, в одном гене – участке ДНК, кодирующем овальбумин курицы, со- держится 7 интронов, в гене сывороточного альбумина крысы – 13 интро- нов. Длина интрона бывает различной – от 200 до 1 тыс. пар нуклеотидов ДНК. Интроны считываются (транскрибируются) одновременно с экзо- нами, поэтому про-мРНК значительно длиннее, чем зрелая м-РНК. В ядре
в про–мРНК специальными ферментами вырезаются интроны, а фрагменты экзона «сращиваются» между собой в строгом порядке. Этот процесс называется «сплайсингом» (сращивание).
В процессе сплайсинга образуется зрелая м-РНК, которая содержит только ту информацию, которая необходима для синтеза соответствующе- го полипептида, т.е. информативную часть структурного гена. Значение и функции интронов до сих пор еще не совсем выявлены, но установлено, что если в ДНК считываются только участки экзонов, зрелая м-РНК не образуется.
13
На специальных генах синтезируется и два других типа молекул РНК
– транспортная и рибосомная, причем для синтеза последней в молекуле ДНК имеется много участков. Кроме того, значительная часть р-РНК син- тезируется на ДНК, содержащейся в ядрышке. В некоторых случаях, на- пример у бактерий, одна молекула м-РНК может синтезироваться на уча- стке ДНК, называемом опероном и включающем несколько генов, обу- словливающих синтез ферментов, которые последовательно влияют на развитие одногопризнака.
Установлено, что по продолжительности существования м-РНК мож- ноподразделитьна:
1)бактериальную, быстро разрушающуюся, что, по-видимому, связано с коротким временем жизни самой бактериальной клетки;
2)м-РНК эмбриональных клеток, лейкоцитов и различных клеток млекопитающих, существующую около 3-4 часов;
3)стабильную, которая функционирует длительное время и наблюдается в специализированных клетках млекопитающих.
Следующий этап биосинтеза – трансляция – происходит в цито-
плазме на рибосомах при участии т-РНК. При этом последовательность оснований в молекуле м-РНК переносится на последовательность аминокислот в полипептидной цепи, образующейся на этой молекуле. Образовавшись в ядре, м-РНК через поры ядерной мембраны
поступает в каналы на эндоплазматической сети или в гиалоплазму и здесь соединяется с рибосомами.
Транспортные РНК синтезируются в ядре, но функционируют в сво- бодном состоянии в цитоплазме клетки. Одна молекула т-РНК содержит 76-85 нуклеотидов и имеет довольно сложную структуру, напоминающую клеверный лист.
Три участка т-РНК имеют особо важное значение:
1)антикодон, состоящий из трех нуклеотидов, определяющий место прикрепления т-РНК к соответствующему комплементарному кодону (м-РНК) на рибосоме;
2)участок, определяющий специфичность т-РНК, способность данной молекулы прикрепляться только к определенной аминокисло-
те;
3) акцепторный участок, к которому прикрепляется аминокисло-
та.
Он одинаков для всех т-РНК и состоит из трех нуклеотидов – Ц-Ц-А. Присоединение аминокислоты к т-РНК предшествует ее активация фер-
14

ментом аминоацил – т-РНК – синтетазой, выделяемый митохондрия- ми. Этот фермент специфичен для каждой аминокислоты. Активирован- ная аминокислота прикрепляется к соответствующей т-РНК и доставляет- ся ею на рибосому. Молекулярный вес т-РНК около 27-28 тыс.
По нуклеотидному составу т-РНК несколько отличается от других ри- бонуклеиновых кислот, т.к., помимо обычных А, Г, Ц и У, в нее входят редкие нуклеотиды (7-8 в каждой т-РНК). Их роль заключается в том, что они повышают сопротивляемость т-РНК действию фермента рибонуклеа- зы, благодаря чему т-РНК существует более длительный период. К на- стоящему времени изучено строение т-РНК для семи разных аминокис- лот.
Центральное место в трансляции принадлежит рибосомам – рибонук- леопротеиновым органоидам цитоплазмы, во множестве в ней присутст- вующим. Размеры рибосом у прокариот в среднем 30 х 30 х 20 нм, у эука- риот – 40 х 40 х 20 нм. Обычно их размеры определяют в единицах седи- ментации (S) – скорости осаждения при центрифугировании в соответст- вующей среде. У бактерии кишечной палочки рибосома имеет величину 80 S и состоит из двух субчастиц, одна из которых имеет константу 30S, вторая 50S и содержит 64% рибосомальной РНК и 36% белка. Участок рибосомы, взаимодействующий с м-РНК, находится на меньшей субъе- динице в части, примыкающей к большей, и связывает при участии рибо- сомальных белков и рибосомной РНК, примерно около 25 оснований мо- лекулы м-РНК. Трансляция начинается с так называемого стартового ко- дона (инициатора синтеза) - А-У-Г-. Когда т-РНК доставляет к рибосоме активированную аминокислоту, ее антикодон соединяется водородными связями с нуклеотидами комплементарного кодона м-РНК. Акцепторный конец т-РНК с соответствующей аминокислотой прикрепляется к рибосо- ме, после первой аминокислоты другая т-РНК доставляет следующую аминокислоту, и таким образом на рибосоме синтезируется полипептид- ная цепь. Молекула м-РНК обычно работает сразуже на нескольких (5-20) рибосомах соединенных в полисомы.
Начало синтеза полипептидной цепи называют инициацией, рост ее элонгацией. Последовательность аминокислот в полипептидной цепи определяется последовательностью кодонов в м-РНК. Синтез полипеп- тидной цепи прекращается, когда на м-РНК появляется один из кодонов – терминаторов – УАА, УАГ или УГА. Окончание синтеза данной поли-
пептидной цепи называется терминацией.
15

Установлено, что в клетках животных полипептидная цепь за одну се- кунду удлиняется на 7 аминокислот, а м-РНК продвигается на рибосоме на 21 нуклеотид. У бактерий этот процесс протекает в 2-3 раза быстрее. Следовательно, синтез первичной структуры белковой молекулы – поли- пептидной цепи – происходит на рибосоме в соответствии с порядком че- редования нуклеотидов в матричной рибонуклеиновой кислоте м-РНК.
1.3.Генетический код. Наиболее неясным в синтезе белков был во- прос о том, как т-РНК находит соответствующий участок м-РНК, к кото- рому должна быть присоединена приносимая ею аминокислота. Это зави- сит от последовательности нуклеотидов в цепи молекулы и-РНК, причем заранее можно было утверждать, что такого рода генетический код не мо- жет состоять из одного или двух нуклеотидов, т.к. последних только четы- ре, сочетаний из двух нуклеотидов 16, а аминокислот 20. Следовательно, если бы код включал даже по два нуклеотида, то это вызвало бы неизбеж- ную путаницу в синтезе определенной белковой молекулы. Отсюда было высказано предположение, что генетический код должен включать не ме- нее трех нуклеотидов, т.е. должен быть триплетным.
Кодом наследственности или генетическим кодом называется процесс перевода триплетной последовательности нуклеотидов молекулы ДНК в последовательность аминокислот в белковой молекуле. Од-
ним из важнейших свойств генетического кода является его колинеар-
ность – четкое соответствие междупоследовательностями кодонов нуклеиновых кислот и аминокислотами полипептидных цепей.
Важное значение для раскрытия генетического кода имели исследова- ния М.Ниренберга и Дж.Маттеи, а затем С.Очоа с сотрудниками, начатые ими в 1961 г. в США. Они разработали метод и экспериментально устано- вили последовательность нуклеотидов в кодонах м-РНК, контролирую- щая местоположение данной аминокислоты в полипептидной цепи.
В бесклеточную среду, содержащую все аминокислоты, рибосомы, т- РНК, АТФ и ферменты, Ниренберг и Маттеи вводили искусственно син- тезированный биополимер типа м-РНК, представляющий собой цепочку одинаковых нуклеотидов – УУУ-УУУ-УУУ… Биополимер кодировал синтез полипептидной цепи, содержащей только одну аминокислоту – фенилаланин; такая цепь называется полифенилаланином. Если м-РНК состояла из кодонов, содержащих нуклеотиды с азотистым основанием цитозин, то синтезируется полипептидная цепь, содержащая аминокисло- тупролин, - полипролин.
16
В дальнейшем исследователи получали искусственные полимеры типа м-РНК, содержавшие, помимо урацила, одно или два других основания А, Г или Ц в разных пропорциях. Они наблюдали, что в зависимости от вто- рого основания и его соотношения с У получаемые полипептиды включа- ли, помимо фенилаланина, какую-нибудь другую аминокислоту или даже две их. Аналогичное явление наблюдалось и при замене урацила А, Г или Ц. Из 20 аминокислот 18 оказались закодированными одним или двумя
основаниями и только две требовали для включения в полипептид три основания. Следовательно, представление о триплетности кода этими
опытами было доказано. Для трех нуклеотидов, обусловливающих включение той или иной аминокислоты в молекулу белка, был предложен термин «кодон» – происходящий от слова «код».
Следующим этапом расшифровки генетического кода было изучение последовательности оснований в кодоне. Ведь первыми исследованиями было установлено лишь то, какие основания входят в состав кодона, но, например, при составе кодона 2ГУ основания в нем могут располагаться в последовательности ГГУ, ГУГ или УГГ, что будет влиять на включение той или иной аминокислоты.
Большие и тонкие исследования в указанном направлении были про- ведены Нирнбергом и его сотрудниками; они использовали для этой цели метод, основанный на том, что первая стадия синтеза белка в рибосомах заключается в присоединении т-РНК, несущих ту или иную аминокисло- ту, к определенному кодону м-РНК. Оказалось, что для такой связи не обязательно нужна целая молекула м-РНК: достаточно использовать со- стоящие их трех оснований соединения, в котором их последовательность легко точно установить.
В результате этих исследований и работ других ученых, устанавли- вавших последовательность оснований в кодоне путем изучения мутаци- онных изменений в структурах белков, последовательность оснований в кодонах уже в 1965 г. была выявлена. Из 64 кодонов, возможных при ком- бинации трех оснований, 61 определяют включение в молекулу полипеп- тида соответствующей аминокислоты, а три определяют конец трансля- ции (своего рода запятые при синтезе белков). К ним не присоединяется ни одна т-РНК, и в синтезе полипептидной цепи они не участвуют.
1.Генетический код вырожденный, т.е. одна иминокислотаможет кодироваться несколькими (от одного до 6) кодонами. Только две аминокислоты кодируются одним триплетом-метионин (АУГ) и триптофан (УГГ).
17
2.Генетический код не перекрывающийся. Нуклеотидная последовательность считывается подряд в одном направлении, триплет за триплетом, т.е. молекула т-РНК, несущая одну аминокислоту, занимает один триплет молекулы м-РНК, вторая соседний и т.д.
3.Генетический код универсален – един для всех организмов (виру-
сов, бактерий, растений, животных и человека), т.е у всех организмов для определенных аминокислот кодоны одинаковы.
Вместе с тем следует отметить, что это не означает, что у всех орга- низмов всегда функционируют все кодоны. Имеют ли они какой-либо «смысл» для данного вида или «бессмысленны», т.е. используются ли они или не используются при синтезе белков, зависит от состава т-РНК, кото- рый у представителей разных видов может быть неодинаковым. Дело в том, что каждому кодону соответствует определенная т-РНК, комплемен- тарная по одному триплету, соответствующему кодону, к которому она и присоединяется, ставя в цепь полипептида соответствующую аминокис- лоту.
Если у данного вида отсутствует т-РНК, соответствующая по своему строению, например, кодону УГГ, то для этого вида он становится «бес- смысленным», т.к. в белковом синтезе не участвует. Наряду с такими «бессмысленными» лишь для определенных видов кодонами, имеется три кодона, которые «бессмысленны» для всех видов УАА, УАГ и УГА. Они играют большую роль при синтезе полипептидной цепи на молекуле м- РНК, включающей несколько генов, являясь своего рода точками, на ко- торых прекращается синтез одной полипептидной цепи, после чего начи- нается синтез другой. Возможно, что эти кодоны имеют значение и для процесса транскрипции, указывая местоначала гена в молекуле ДНК.
4.Код триплетный. Местоположение каждой аминокислоты кодируется сочетанием строго определенных трех нуклеотидов в м- РНК, образующих один специфический кодон.
5.Кодон АУГ, находящийся в начале и-РНК является инициатором синтеза полипептидной цепи. Если данный кодон находится в середине м-РНК, то он кодирует аминокислоту метионин.
6.Кодоны УАГ («амбер»), УАА («охра») и УГА («опал») являются терминаторами (стоп-сигналами) синтеза. Когда считывание генетической информации в м-РНК доходит до одного из этих кодонов, дальнейший синтез прекращается и полипептидная цепь отделяется от рибосомы.
18
Следовательно, в каждой клетке в молекулах ДНК закодирована вся генетическая информация, которая может быть реализована в онтогенезе через биосинтез в виде биохимических процессов, физиологических свойств
иморфологических признаков.
1.4.Регуляция активности генов. Изучение химического состава кле- ток, полученных из разных тканей одного многоклеточного организма, показывает, что каждая из них содержит разный, относительнонебольшой набор белковых молекул, хотя все они имеют одинаковый набор хромо- сом, и, следовательно, единую генетическую информацию. Так, у бакте- рии E.coli в одной клетке в различные периоды ее жизнедеятельности комплекс ферментов бывает разный. Все это дало основание предполо- жить, что в клетке имеется механизм, регулирующий активность генов, определяющий, какие гены в данный момент должны быть активными и
каким следует находится в неактивном, репрессированном состоянии.
Механизм регуляции генетического кода был открыт французскими учеными Ф.Жакобом и Ж.Моно в 1961 г. на бактериях E.coli и получил название механизма индукции – репрессии. Было установлено, что синтез соответствующих белков – ферментов индуцируется веществом, служащим субстратом для данного фермента и необходимым для нормальной жизнедеятельности клетки.
Так, например, для нормальной жизнедеятельности E.coli необходим молочный сахар (лактоза) и в ее геноме содержатся гены, контролирую- щие синтез ферментов, гидролизующих лактозу до простых соединений. Если среда, в которой находятся бактерии, лактозы не содержит, эти гены пребывают в репрессированном состоянии и не функционируют. Внесен- ная в среду лактоза будет тем индуктором, который включает в работу данные гены, и в клетке начинается синтез ферментов, гидролизующих лактозу до более простых соединений. После удаления лактозы из среды синтез этих ферментов прекращается.
Роль репрессора может выполнять и вещество, синтезируемое в клет- ке, если содержание его превышает норму. Например, если синтезируются нуклеотиды, аминокислоты и другие вещества и содержание их превыша- ет количество, необходимое данной клетке, каждое из них может быть репрессором и подавлять работу генов, синтезирующих ферменты, необ- ходимыедля данногобиохимического процесса.
Механизм индукции – репрессии обеспечивает включение (индук- цию) в работу тех генов, которые синтезируют необходимые на данном этапе жизнедеятельности клетки ферменты. Работа генов прекращается
19
(репрессируется), когда деградируемый данными ферментами субстрат
израсходован или когда синтезируемое данными ферментами вещество находится в избытке.
По теории Жакоба и Моно, гены, влияющие на синтез определенного фермента или белка, расположены в молекуле ДНК последовательно друг за другом в порядке их влияния на ход реакции синтеза. Такая группа ге- нов называется опероном, а гены, непосредственно кодирующие синтез соответствующих ферментов, называют структурными генами. В на- чале каждого оперона находится специальный ген – ген оператор. На структурных генах одного оперона обычно образуется одна м-РНК, и эти гены бывают одновременно активны или неактивны. Как правило, струк- турные гены в опероне находятся в состоянии репрессии.
Включение в работу оперона или выключение его осуществляет уча- сток ДНК, получивший название гена – регулятора. Ген-регулятор рас- положен на особом участке молекулы ДНК и кодирует синтез белка – ре- прессора, выключающего работу оперона. Соединяясь с геном – операто- ром, репрессор замыкает двойную цепь молекулы ДНК, прекращая этим синтез м-РНК на структурных генах и образование первого из ферментов, необходимого для хода реакции синтеза.
Работой структурных генов управляют находящиеся в опероне гены, не имеющие кодирующих функций. Их называют акцепторными генами. Система акцепторных и структурных генов образуют один оперон. Оперон состоит из нескольких участков ДНК, каждый из которых принимает активное участие в транскрипции м-РНК: промотора, оператора, спейсера, структурных генов и терминатора.
Промотор – участок ДНК, опознаваемый ферментом РНК – полимеразой, обеспечивающим синтез м-РНК.Ему в опероне предшествует участок ДНК, к которому присоединяется Сар-белок – белок активатор. Эти два участка ДНК состоят из 85 нуклеотидных пар. После промотора в опероне размещается ген – оператор, состоящий из 21 нуклеотидной пары. С ним обычно и бывает связан белок – репрессор, вырабатываемый геном-регулятором. За геном-оператором располагается спейсер (space –промежуток). Спейсеры – неинформативные участки молекулы ДНК различной длины (иногда до 20000 пар оснований), которые, по видимому, принимают участие в регулировании процессатранскрипции соседнего гена.
После спейсера в опероне располагаются структурные гены, число ко- торых может быть различным. У кишечной палочки lac-оперон содержит
20