

І, іі, ііі класів на 6-й хромосомі людини
Між доменами α1 і α2 утворюється своєрідна щілина (активний центр), в яку входять епітопи антигену при імунній відповіді, причому в щілину, як правило, вміщується 8-10 амінокислотних залишків.
Рис.
7.2. Структура антигенів МНС класів І і
ІІ
Позначення:
α і β – домени пептидних ланцюгів; ВЛ
– вуглеводні ланцюги; ЦПМ – цитоплазматична
мембрана; βм –
β-мікроглобулін.
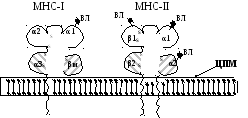
Антигени МНС класу ІІ (МНС-ІІ) присутні лише на антигенпрезентуючих клітинах: макрофагах, дендритних клітинах, В-лімфоцитах, а у деяких видів також на активованих Т-лімфоцитах та судинному ендотелії. Ці антигени кодуються генними локусами ділянки D (або Ia – імуноасоційована), що розташована у людини на шостій хромосомі поряд з HLA-А, HLA-В, HLA-С. Ділянка D, у свою чергу, поділяється на 3 генних локуси: HLA-DP, HLA-DQ, HLA-DR.
Ці гени кодують 2 поліпептидних ланцюги – α і β, молекулярна маса яких становить відповідно 35 та 25 кД. Обидва мають цитоплазматичний фрагмент з С-кінця, гідрофобну ділянку, заякорену у мембрану, і по два петлеподібних домени, що розташовані над мембраною. Деякі з них з’єднано дисульфідними зв’язками. Домени α1іβ1є гіперваріабельними і високоспецифічними для зв’язування епітопів антигенів.За специфічністю вони незначно поступаються антитілам. Ймовірно, що вони теж кодуються багатьма алелями. Між доменами α1 і β1 утворюється щілина активного центру, яка може приєднуватиепітопи антигенів. МНС-ІІ здатні зв’язувати більш великі епітопи, ніж МНС-І (13-17 амінокислот, а можливо, 30 і більше). Як вже зазначалося, антигени МНС-ІІ присутні тільки на імуноцитах. У моноцитах, макрофагах, В-лімфоцитах проходить або фагоцитування, або поглинання антигену шляхом ендоцитозу, і він фрагментується за допомогою лізосомальних ферментів. Великі епітопи і деякі фрагменти з’єднуються з антигенами MHC-II і виводяться на поверхню ЦПМ. Такий комплекс розпізнається Т-хелперами 2 за допомогою TCR і включається гуморальна імунна відповідь.
Надзвичайно важливу роль відіграє головний комплекс гістосумісності у реалізації імунної відповіді, а саме у розпізнаванні антигенів. Але різні за складом і природою антигени експресуються на поверхню клітин у комплексі або з МНС-І, або з МНС-ІІ.
Звичайно з’єднання епітопа антигену з поліпептидними ланцюгами МНС відбувається всередині антигенпрезентуючої або ураженої клітини в порожнині ендоплазматичного ретикулюма (ЕПР). На шорсткуватому ЕПР йде синтез антигенів MHC, а всередині його формується комплекс з епітопом, що потім попадає в пухирці апарата Гольджі, а з них процесується на поверхню цитоплазматичної мембрани. У такому вигляді антигени розпізнаються Т-хелперами і Т-ефекторами (ЦТЛ – цитотоксичними лімфоцитами), що веде до включення гуморальної або клітинної імунної відповіді.
В останні роки висловлюється припущення, що клітинна імунна відповідь опосередковується в основному Тh-1, а гуморальна – Тh-2. Т-хелпери взагалі мають на своїй поверхні специфічний TCR-рецептор, що бере участь у розпізнаванні комплексу “МНС – антиген”. Суть його полягає в тому, що Т-хелпери впізнають не тільки МНС, але й адсорбований на ньому епітоп чужерідного антигену, тобто відбувається розпізнавання “зміненого свого” (подвійне розпізнавання антигенів).
Виявилося, що антигени внутрішньоклітинних паразитів (вірусів, гонококів, хламідій, токсоплазм тощо), а також ракові і трансплантаційні антигени експресовані на мембрані уражених клітин в комплексі з МНС-І. Такий комплекс запускає клітинну імунну відповідь, тому що МНС-І розпізнаються Тh-1 і Т-ефекторами (Т-кілерами).
Антигени, “перетравлені” макрофагами (бактерії, токсини, яди та всі розчинні антигени) експресуються на мембрани фагоцитуючих клітин (іноді В-лімфоцитів) в комплексі з МНС-ІІ. Такий комплекс розпізнається Т-хелперами (в основному Тh-2), внаслідок чого вмикається гуморальна імунна відповідь. Але розвиток клітинної, або гуморальної відповіді залежить значною мірою ще від того, які цитокіни утворюються імуноцитами під впливом антигену.
Антигени МНС класу ІІІ (МНС-ІІІ)включають розчинні сироваткові фактори, які не фіксуються на клітинах, а циркулюють у крові. Це такі фактори, як: С2-, С4а- і С4b-компоненти комплементу, а також Вf -компонент пропердину. Гени, що кодують ці фактори, також розташовані у 6 хромосомі між генними локусами МНС-І і МНС-ІІ.
Антигени головного комплексу гістосумісності виконують в організмі важливі функції:
беруть участь в імунному розпізнаванні антигенів;
індукують бласттрансформацію Т- і В-лімфоцитів;
стимулюють утворення антитіл;
беруть участь у реакції лімфолізу (руйнування лімфоцитів при контакті з антигеном і активацією комплемента);
контролюють силу імунної відповіді і сприяють її пригніченню завдяки наявності двох додаткових генів поблизу D-ділянки:Ir(immuneresponse– ген, що регулює силу імунної відповіді) іIs(від англ.immune supрress– ген, що пригнічує імунну відповідь);
опосередковують реакції відторгнення трансплантату;
беруть участь у реакції “трансплатат - проти хазяїна”, яка виникає при пересаджуванні донорського кісткового мозку і супроводжується імунною відповіддю проти реципієнта.
Генетичними дослідженнями було встановлено певний зв’язок між наявністю конкретних антигенів МНС і схильністю до цілого ряду захворювань. Так, вроджені аномалії серця частіше виявляються у людей з антигеном HLA-А2; важке захворювання міастенія обумовлюється наявністюHLA-В8, такий же антиген сприяє розвитку системної червоної вівчанки; у 90% людей з HLA-В27 розвивається анкілозуючий спондиліт. HLA-DRW3– часто виявляється у людей, схильних до герпесного дерматиту. ЗHLA-А3пов’язують імунологічну дефектність у відношенні до деяких вірусів.