

Дуус / Топический диагноз в неврологии
.PDF28 1 Система чувствительности
N. maxillaris

 Сегментарная иннервация
Сегментарная иннервация 
 Hansen*Schliack).
Hansen*Schliack).
чем рост спинного мозга. Корешки, на5 |
ся на уровне третьего поясничного по5 |
|||
правляясь к свои межпозвонковым от5 |
звонка. |
|||
верстиям, идут косо вниз и становятся |
За исключением сегментарного от5 |
|||
все длиннеее и длиннее по направлению |
хождения нервных корешков, в самом |
|||
к концу спинного мозга |
|
спинном мозге не обнаруживается мор5 |
||
|
фологических признаков метамерного |
|||
ного мозга), который обычно находится |
||||
деления (см. рис. 1.22). Несоответствие |
||||
на уровне 25го поясничного |
позвонка. |
|||
между локализацией сегментов спинно5 |
||||
Ниже этого уровня |
субарахноидальное |
|||
го мозга и соответствующих позвонков, |
||||
пространство, имеющее здесь вид сум5 |
||||
увеличивающееся по мере приближения |
||||
ки, содержит лишь передние и задние |
||||
к конусу, должно учитываться при оп5 |
||||
корешки, которые |
образуют |
конский |
ределении уровня поражения спинного |
|
хвост (рис. 2.34). В редких случаях ко5 |
мозга. |
|||
нус спинного мозга может заканчивать5 |
Корешки сегментов спинно5 |
|||
Спинной мозг и периферическая иннервация |
29 |
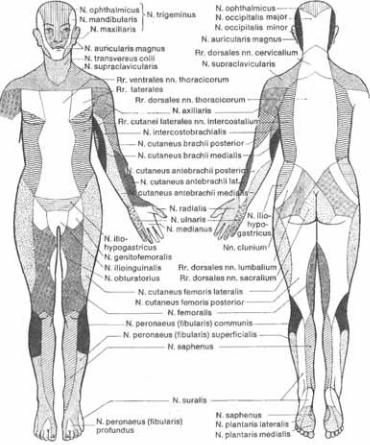
 1.29. Распределение кожной чувствительности соответственно периферическим нервам.
1.29. Распределение кожной чувствительности соответственно периферическим нервам.
го мозга покидают позвоночный канал через межпозвонковые отверстия, рас5 положенные на уровне верхнего (ораль5 ного) края тела каждого позвонка. По5 скольку шейный отдел спинного мозга имеет на один сегмент больше по срав5 нению с числом шейных позвонков, то корешок 85го сегмента покидает канал через межпозвонковое отверстие, распо5 ложенное между 75м шейным и 15м грудным позвонками. С этого уровня и ниже нервные корешки проходят через межпозвонковое отверстие на уровне нижнего края соответствующего по5 звонка.
Между С4 и ТЫ, а также между L2 и S3 сегментами диаметр спинного мозга увеличивается. Шейное и пояснич$ ное утолщения возникают в связи с тем, что корешки нижней половины шейно5 го отдела спинного мозга дают начало плечевому сплетению, иннервирующе5 му верхние конечности, а корешки люм5  отдела образуют люмбо5 сакральное сплетение, иннервирующее нижние конечности (рис. 122).
отдела образуют люмбо5 сакральное сплетение, иннервирующее нижние конечности (рис. 122).
При образовании сплетений каждая пара корешков отдает ветви к несколь5 ким различным периферическим нер5 вам; иными словами, каждый перифе5

30 1 Система чувствительности
 spinalis (Tractusspinothalamicus
spinalis (Tractusspinothalamicus
 et lateralis)
et lateralis)
Tractus spinothalamicus lateralis
 spinothalamicus lateralis
spinothalamicus lateralis
Nucleus sensorius principalis n. 
Nucleus et tractus spinalis n. 
 1.30. Клиническиесиндромы поражения проводников чувстви* тельности на уровнях, отмеченных красными линиями и обозначен* ных буквами латинского алфавита (подробности в тексте).
1.30. Клиническиесиндромы поражения проводников чувстви* тельности на уровнях, отмеченных красными линиями и обозначен* ных буквами латинского алфавита (подробности в тексте).
рический нерв образован волокнами не5 |
дерматомами, или дерматомными |
скольких корешков, выходящих из при5 |
зонами (рис. 1.26). |
лежащих друг к другу сегментов спин5 |
Дерматомов имеется столько, сколь5 |
ного мозга (рис. 1.23, 124 и 1.25). До5 |
ко и сегментов спинного мозга, отда5 |
стигнув периферии, афферентные во5 |
ющих нервные корешки. На рис. 1.28 |
локна вновь соединяются соответствен5 |
показаны все дерматомы тела (вид спе5 |
но их первоначальной принадлежности |
реди и сзади). Лучше всего метамерная |
задним корешкам и иннервируют оп5 |
организация дерматомов видна на при5 |
ределенные сегменты кожи, называе5 |
мере грудного отдела. Все дерматомы |

Спинной мозг и периферическая иннервация |
31 |
частично перекрывают друг друга (рис. 126), поэтому выпадение функции од5 ного отдельного корешка обнаружить бывает трудно. Лишь выпадение функ5 ции нескольких смежных корешков приводит к сегментарному нарушению чувствительности. Тот факт, что каждый дерматом соответствует определенному сегменту спинного мозга, имеет боль5 шое диагностическое значение при оп5 ределении уровня поражения спинного мозга. Рис.  приводится для того, чтобы легче было понять и запомнить сегментарную иннервацию тела (шей5 ные, грудные, поясничные и крестцовые дерматомы).
приводится для того, чтобы легче было понять и запомнить сегментарную иннервацию тела (шей5 ные, грудные, поясничные и крестцовые дерматомы).
Безусловно, при повреждении части сплетения или отдельных перифериче5 ских нервов картина нарушения чувст5 вительности будет отличаться от той, которая наблюдается при повреждении корешков. Вовлечение в патологический процесс сплетения приводит к преиму5 щественному нарушению двигательных функций в виде типичных синдромов, которые будут описаны в ниже в главе, посвященной двигательным функциям.
Необходимо повторить еще раз, что волокна, образующие периферический нерв, происходят из различных кореш5 ков. При повреждении нерва волокна, иннервирующие часть дерматома, не могут объединиться с волокнами, ин5 нервирующими другую его часть, по5 скольку они идут в составе различных нервов. Следовательно, нарушение чув5 ствительности, вызванное повреждени5 ем периферического нерва, полностью отличается от нарушения чувствитель5 ности, обусловленного поражением ко$ решка. Перекрытие зон иннервации со5 седних нервов значительно меньше, чем перекрытие зон корешковой иннерва5 ции, что существенно облегчает выяв5 ление чувствительных нарушений при поражении нервных стволов.
В дерматомах зоны перекрытия так5 тильной чувствительности больше, чем
болевой. Таким образом, при повреж5 дении одного или двух корешков сни5 жение болевой и температурной чувст5 вительности обнаружить легче, чем сни5 жение тактильной чувствительности. Это необходимо учитывать для выявле5 ния гипалгезии или анальгезии при по5 дозрении на корешковое повреждение.
При повреждении периферического нерва зона гипестезии существенно больше зоны гипалгезии. Поэтому ги5 пестезия выявляется значительно рань5 ше. Иногда бывает трудно отличить чув5 ствительные нарушения, вызванные по5 ражением С8 корешка, от нарушений чувствительности, обусловленных по5 вреждением локтевого нерва, а также нарушения чувствительности при пора5 жении  корешков от нарушений чувствительности,
корешков от нарушений чувствительности,  в
в  с патологией малоберцового нерва. Зоны чувствительных нарушений в рассмот5 ренных случаях практически идентич5 ны, как видно из сравнения рисунков 1.28 и 129.
с патологией малоберцового нерва. Зоны чувствительных нарушений в рассмот5 ренных случаях практически идентич5 ны, как видно из сравнения рисунков 1.28 и 129.
Каждый периферический чувстви5 тельный нерв имеет четко очерченную зону иннервации, что позволяет выяв5 лять поражение нерва путем тщатель5 ного обследования. Например, можно обнаружить, что зона дизестезии соот5 ветствует зоне иннервации наружного кожного нерва бедрй и что именно этот нерв ответственен за 
Синдромы поражения проводников
 нарушения чувствительно5 сти различаются в зависимости от уров5 ня поражения чувствительных провод5 никовых путей. На рис. 1.30 показаны десять различных локализаций возмож5 ного поражения, обозначенных красны5 ми поперечными линиями и помечен5 ные буквами латинского алфавита (от
нарушения чувствительно5 сти различаются в зависимости от уров5 ня поражения чувствительных провод5 никовых путей. На рис. 1.30 показаны десять различных локализаций возмож5 ного поражения, обозначенных красны5 ми поперечными линиями и помечен5 ные буквами латинского алфавита (от  до
до 
32 1 Система чувствительности
Локализация а или  корковое или подкорковое поражение сенсомоторной зоны руки (а) или ноги
корковое или подкорковое поражение сенсомоторной зоны руки (а) или ноги  вызывает парестезии (покалывание, мурашки и т. д.) и онемение в соответствующей конечности на противоположной сторо5 не, более выраженные в дистальных от5 делах. Парестезии могут возникать в форме фокальных сенсорных припад5 ков. При вовлечении двигательных зон коры
вызывает парестезии (покалывание, мурашки и т. д.) и онемение в соответствующей конечности на противоположной сторо5 не, более выраженные в дистальных от5 делах. Парестезии могут возникать в форме фокальных сенсорных припад5 ков. При вовлечении двигательных зон коры  также двигательные фокальные припадки (Джексоновские атаки).
также двигательные фокальные припадки (Джексоновские атаки).
Локализация с: поражение, вовлека5 ющее все чувствительные пути непо5 средственно перед их входом в 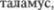 вызывает нарушение всех видов чувст5 вительности в противоположной поло5 вине тела.
вызывает нарушение всех видов чувст5 вительности в противоположной поло5 вине тела.
Локализация d: если повреждаются все чувствительные пути за исключе5 нием проводников болевой и темпера5 турной чувствительности, то гипестезия наступает на противоположной стороне лица и  при этом болевая и температурная чувствительности сохра5 няются.
при этом болевая и температурная чувствительности сохра5 няются.
Локализация е: если зона поражения расположена в стволе мозга и ограни5 чивается областью петли тройничного нерва и латерального спинно5таламиче5 ского пути, то на противоположной сто5 роне тела и туловища нарушается бо5 левая и температурная чувствитель5 ность, а все остальные виды чувстви5 тельности остаются сохранными.
Локализация f: при поражении ме5 диальной петли и переднего спинно5та5 ламического пути на противоположной стороне тела нарушаются все виды чув5 ствительности, кроме болевой и темпе5 ратурной.
Локализация g: повреждение ядра и спинального пути тройничного нерва и латерального спинноталамического пу5 ти приводит к нарушению болевой и температурной чувствительности на ип5  стороне лица и контрла5 теральной стороне туловища.
стороне лица и контрла5 теральной стороне туловища.
Локализация h: повреждение задних канатиков вызывает нарушение чувства позы, вибрационной, дискриминацион5 ной и других видов глубокой чувстви5 тельности, приводящее к развитию атаксии на стороне поражения.
Локализация i: повреждение задних рогов спинного мозга ведет к утрате болевой и температурной чувствитель5 ности на стороне поражения; все ос5 тальные виды чувствительности оста5 ются сохранными (диссоциированное расстройство чувствительности).
Локализация к: повреждение не5 скольких соседних задних корешков со5 провождается корешковой парестезией и болью, а также снижением или утра5 той всех видов чувствительности в со5 ответствующем сегменте тела. Если по5 вреждаются корешки, интернирующие •руку или ногу, то, кроме того, наблю5 дается гипотония или атония, арефлек5 сия и атаксия.
2 Двигательная система
Произвольные движения мышц связа5 ны с длинными нервными волокнами, которые начинаются от корковых ней5 ронов и спускаются к клеткам передних рогов спинного мозга. Эти волокна фор5 мируют 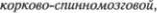 или
или
пирамидный путь, tractus
seu 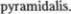 Они являются аксонами нервных клеток, расположенных в дви5 гательной зоне коры, в
Они являются аксонами нервных клеток, расположенных в дви5 гательной зоне коры, в 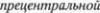
извилине,  praecentralis, преимуще5 ственно в 45м цитоархитектоническом поле Бродмана (рис. 2.1). Это поле пред5 ставляет сравнительно узкую полосу, распространяющуюся вдоль централь5 ной борозды от латеральной, или силь5 виевой, борозды до верхнего края полу5 шария и затем по передней части па5 рацентральной дольки на медиальной поверхности полушария. Оно лежит на5 против чувствительной коры постцент5 ральной извилины. Нейроны,
praecentralis, преимуще5 ственно в 45м цитоархитектоническом поле Бродмана (рис. 2.1). Это поле пред5 ставляет сравнительно узкую полосу, распространяющуюся вдоль централь5 ной борозды от латеральной, или силь5 виевой, борозды до верхнего края полу5 шария и затем по передней части па5 рацентральной дольки на медиальной поверхности полушария. Оно лежит на5 против чувствительной коры постцент5 ральной извилины. Нейроны,  рующие глотку и гортань, расположены в его нижней части, вблизи
рующие глотку и гортань, расположены в его нижней части, вблизи 
борозды. Затем в восходящем порядке следуют нейроны к мышцам лица,  туловища и ног (рис. 22). Такая сома5 тотопическая проекция соответствует стоящему на голове человеку. Схема расположения человека в чувствитель5 ной коре постцентральной извилины приведена ранее (рис. 1.20).
туловища и ног (рис. 22). Такая сома5 тотопическая проекция соответствует стоящему на голове человеку. Схема расположения человека в чувствитель5 ной коре постцентральной извилины приведена ранее (рис. 1.20).
Мотонейроны расположены не толь5 ко в поле 4; они также встречаются в соседних кортикальных полях. Однако основная их масса занимает 5 корти5
 слой 45го цитоархитектониче5 ского поля. Они обеспечивают отдель5 ные точные целенаправленные движе5 ния. Эти нейроны включают гигантские пирамидные клетки Беца, дающие ак5 соны с толстой миелиновой оболочкой (рис. 2.3). Такие быстропроводящие ак5 соны составляют лишь от
слой 45го цитоархитектониче5 ского поля. Они обеспечивают отдель5 ные точные целенаправленные движе5 ния. Эти нейроны включают гигантские пирамидные клетки Беца, дающие ак5 соны с толстой миелиновой оболочкой (рис. 2.3). Такие быстропроводящие ак5 соны составляют лишь от 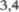 до
до 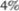 всех волокон пирамидного пути. Основ5 ная часть волокон пирамидного тракта начинается от мелких пирамидных, или веретенообразных, клеток в двигатель5 ных полях 4 и 6. Поле 4 дает около
всех волокон пирамидного пути. Основ5 ная часть волокон пирамидного тракта начинается от мелких пирамидных, или веретенообразных, клеток в двигатель5 ных полях 4 и 6. Поле 4 дает около  волокон пирамидного тракта, ос5 тальные начинаются в других полях сенсомоторной области (см. рис. 2.1).
волокон пирамидного тракта, ос5 тальные начинаются в других полях сенсомоторной области (см. рис. 2.1).
Мотонейроны 4 поля контролируют тонкие произвольные движения скелет5 ных мышц противоположной полови5
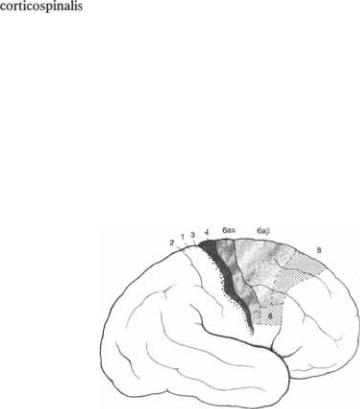
Abb. 2.1
 2.1
2.1 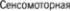 область с первичным моторным полем 4 в прецентральной извилине и премоторными полями 6 и 8.
область с первичным моторным полем 4 в прецентральной извилине и премоторными полями 6 и 8.
34 2 Двигательная система
Рис. 2.2 Пирамидный путь. Пройдя через  венец, его волокна сходятся вместе в области заднего бедра внутренней капсулы в
венец, его волокна сходятся вместе в области заднего бедра внутренней капсулы в  топическом порядке.
топическом порядке.
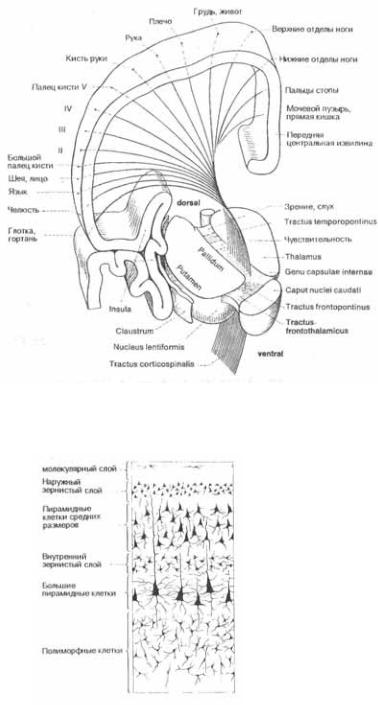
Рис. 2.3 Цитоархитектура двигательной коры (краситель  Крупные клетки формируют пирамидный путь.
Крупные клетки формируют пирамидный путь.
ны тела, поскольку большинство воло5 кон пирамидного тракта переходит на противоположную сторону в нижней ча5 сти продолговатого мозга (рис. 2.4). Стимуляция поля 4 индуцирует общие движения отдельных мышц, в то время как стимуляция поля 6 вызывает более сложные движения, такие, как движения всей руки или ноги.
Импульсы пирамидных клеток дви5 гательной коры идут по двум путям, заключенным в ростральной части пи5 рамидного тракта. Один из них — кор$  путь, tractus corticonuclearis (corticobulbaris), который заканчивается на ядрах черепных двигательных нервов
путь, tractus corticonuclearis (corticobulbaris), который заканчивается на ядрах черепных двигательных нервов
в стволе мозга. Другой — 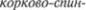
номозговой путь, tractus corticospinalis, представляющий значительно более толстый пучок. Он оканчивается в пе5 редних рогах спинного мозга на вста5

или пирамидный путь |
35 |
Передняя 
Ножка 
|
Medulla |
Decussatio |
|
Tractus |
— |
anterior (прямой) |
(перекрещенный) |
Abb. 2.4
 2.4 Ход пирамидных путей.
2.4 Ход пирамидных путей.
 нейронах, которые, в свою оче5 редь, синаптически связаны с больши5 ми мотонейронами передних рогов. Эти нейроны передают импульсы по перед5 ним корешкам спинного мозга и пери5 ферическим нервам к двигательным концевым пластинкам скелетных мышц (рис. 2.4).
нейронах, которые, в свою оче5 редь, синаптически связаны с больши5 ми мотонейронами передних рогов. Эти нейроны передают импульсы по перед5 ним корешкам спинного мозга и пери5 ферическим нервам к двигательным концевым пластинкам скелетных мышц (рис. 2.4).
Корково*спинномозговой пирамидный путь
Покинув двигательную кору, волокна корково5спинномозгового пути прохо5 дят через белое вещество в составе  чистого венца и сходятся в области зад5 него бедра внутренней капсулы. В очень
чистого венца и сходятся в области зад5 него бедра внутренней капсулы. В очень
4 
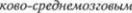
36 2 Двигательная система
компактном виде они проходят в сома5 тотопическом порядке внутреннюю кап5 сулу и вступают в среднюю часть ножек мозга. Здесь они представляют из себя компактный пучок, который опускается
вцентр основания каждой из половин моста, будучи окруженным множеством нейронов ядер моста и волокнами раз5 личных систем. На уровне перехода моста в продолговатый мозг пирамид5 ные пути становятся заметными сна5 ружи и формируют удлиненные пере5 вернутые пирамиды по обе стороны от передней средней линии — отсюда и происходит название — пирамидные пути. В нижней части продолговатого мозга  волокон каждого пира5 мидного тракта переходят на противо5 положную сторону в перекресте пира5 мид и образуют латеральный корково$
волокон каждого пира5 мидного тракта переходят на противо5 положную сторону в перекресте пира5 мид и образуют латеральный корково$
 путь, tractus
путь, tractus 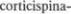

 Оставшиеся неперекрещен5 ными волокна продолжают спускаться
Оставшиеся неперекрещен5 ными волокна продолжают спускаться
впередних канатиках, формируя перед5 ний корково5спинномозговой путь,
tractus  anterior. Его волок5 на переходят на противоположную сто5 рону на сегментарных уровнях в составе передней белой спайки спинного мозга (рис. 2.6). В шейных и грудных сег5 ментах спинного мозга некоторые во5 локна, возможно, соединяются с клет5 ками переднего рога своей стороны;
anterior. Его волок5 на переходят на противоположную сто5 рону на сегментарных уровнях в составе передней белой спайки спинного мозга (рис. 2.6). В шейных и грудных сег5 ментах спинного мозга некоторые во5 локна, возможно, соединяются с клет5 ками переднего рога своей стороны;  ким образом, мышцы шеи и туловища получают корковую иннервацию с обеих сторон.
ким образом, мышцы шеи и туловища получают корковую иннервацию с обеих сторон.
Те волокна, которые пересеклись на уровне перекреста пирамид, спускаются в виде латерального корково5спинно5 мозгового пути в боковом канатике спинного мога, становясь все тоньше и тоньше по мере приближения к пояс5 ничному отделу, так как отдают по пути ответвления. Приблизительно  во5 локон переключаются на вставочные нейроны, которые в свою очередь об5 разуют синапсы с большими альфа5мо5 тонейронами и гамма5мотонейронами
во5 локон переключаются на вставочные нейроны, которые в свою очередь об5 разуют синапсы с большими альфа5мо5 тонейронами и гамма5мотонейронами
передних рогов спинного мозга (рис. 1.10 и 2.4).
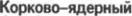 или
или  бульбарный путь
бульбарный путь
Волокна, образующие корково5ядерный путь, покидают ростральный  ный путь на уровне среднего мозга и
ный путь на уровне среднего мозга и
перемещаются несколько дорсальнее, как показано на рис. 2.4, 3.49а и 350а. По пути к ядрам двигательных череп5 ных нервов часть этих волокон перехо5 дит на противоположную сторону, а часть остается неперекрещенной (дета5 ли см. на срезах с ядрами черепных нервов). Пути подходят к ядрам тех нервов, которые обеспечивают произ5 вольную иннервацию лицевой и ораль5 ной мускулатуры: тройничного (V), ли5 цевого (VII),  (IX), блуждающего (X), добавочного (XI) и подъязычного (XII) нервов.
(IX), блуждающего (X), добавочного (XI) и подъязычного (XII) нервов.
Заслуживает внимания и другой пу5 чок. Он начинается не в передней цент5 ральной извилине, а в глазной области 8 поля Бродмана (рис. 2.4 и 2.5). Его импульсы обеспечивают содружествен5 ные движения  Он называется кор$
Он называется кор$
путем, tractus 
 но утвердилось мне5 ние, что в него входит и часть корко5 во5ядерного пути. После того, как во5 локна этого пучка покидают поле 8, они присоединяются к волокнам пирамид5 ного пути в области лучистого венца. Затем они проходят более вентрально через заднюю ножку внутренней капсу5 лы, а далее поворачивают в каудальном направлении по пути к ядрам черепных нервов, обеспечивающих движения глазных яблок: глазодвигательного (III), блокового (IV) и отводящего (VI). Импульсы из поля 8 не ответственны за иннервацию отдельных глазных мышц. Они действуют синергично, обеспечивая содружественные движения
но утвердилось мне5 ние, что в него входит и часть корко5 во5ядерного пути. После того, как во5 локна этого пучка покидают поле 8, они присоединяются к волокнам пирамид5 ного пути в области лучистого венца. Затем они проходят более вентрально через заднюю ножку внутренней капсу5 лы, а далее поворачивают в каудальном направлении по пути к ядрам черепных нервов, обеспечивающих движения глазных яблок: глазодвигательного (III), блокового (IV) и отводящего (VI). Импульсы из поля 8 не ответственны за иннервацию отдельных глазных мышц. Они действуют синергично, обеспечивая содружественные движения

Корково*ядерный или корково*бульбарный путь |
37 |
 2.5 Экстрапирамидные пути.
2.5 Экстрапирамидные пути.
глаз в противоположную сторону  дружественное
дружественное  Не ясно, где
Не ясно, где  волокна корково5 среднемозгового пути. Известно только, что они не образуют прямых синапти5
волокна корково5 среднемозгового пути. Известно только, что они не образуют прямых синапти5  связей с нейронами ядра гла5 зодвигательного нерва (см. срез с изо5
связей с нейронами ядра гла5 зодвигательного нерва (см. срез с изо5
бражением черепных нервов). Для того, чтобы понять работу периферических мотонейронов в стволе мозга и спинном  необходимо рассмотреть также и другие двигательные пути. Они входят в состав экстрапирамидной
необходимо рассмотреть также и другие двигательные пути. Они входят в состав экстрапирамидной  ной системы.
ной системы.