

биология 10-11
.pdf88 |
Глава 4. Размножение организмов |
размножения |
Митоз |
Период |
|
Период роста |
|
Период созревания |
Мейоз |
СПЕРМАТОГЕНЕЗ |
ОВОГЕНЕЗ |
Первое деление мейоза
Второе деление мейоза
Оплодотво- |
Полярные тельца |
рение |
Яйцеклетка
Схема развития сперматозоидов и яйцеклетки Зигота (сперматогенез и овогенез)
приспосабливаться к изменяющимся внешним условиям. В уве личении разнообразия индивидуальной наследственной инфор мации состоит главное преимущество полового размножения, его основное биологическое значение.
Для большинства организмов характерно перекрестное оплодотворение. Широко распространено самооплодотворение только у растений (путем самоопыления), однако у многих обоеполых растений имеются особенности, исключающие само оплодотворение. Тычинки и пестики обоеполых цветков созрева ют не одновременно, поэтому происходит именно перекрестное опыление разных особей. Среди животных самооплодотворение встречается только у гермафродитных форм (у одной особи име ются мужские и женские половые органы), например, у ряда пиявок, брюхоногих моллюсков, некоторых рыб.
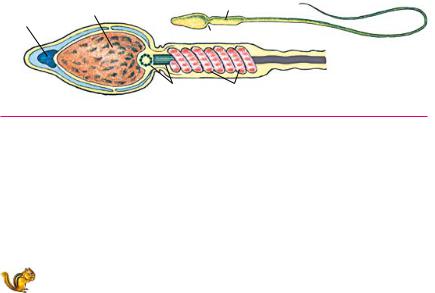
Развитие половых клеток. Формирование половых клеток (гаметогенез) происходит в половых железах. Развитие жен

§ 20. Способы размножения организмов |
89 |
ских гамет (яйцеклеток) происходит в яичниках и носит на звание овогенеза (<лат. ovum яйцо + genesis происхождение). Мужские гаметы (сперматозоиды) формируются в семенниках в процессе сперматогенеза (<греч. spermatos семя). Гаметоге нез происходит последовательно в трех зонах: размножения, роста и созревания. Соответственно выделяют и три периода развития гамет.
В начальный период размножения будущие половые клет ки имеют диплоидный набор хромосом и делятся посредством митоза. Особенно интенсивно размножаются мужские клетки млекопитающих. У мужских особей гаметы образуются прак тически всю жизнь. Формирование яйцеклеток млекопитающих происходит только в эмбриональный период, далее они сохра няются в состоянии покоя.
Попадая в зону роста, будущие половые клетки уже не делятся, а только растут. Предшественники мужских гамет вы растают не слишком сильно, а яйцеклетки увеличивают свои размеры в сотни и миллионы раз (диаметр яйцеклетки кисте перой рыбы латимерии может достигать 9 см). После того как клетки вырастают до размеров взрослых половых клеток, они попадают в зону созревания, в которой происходит мейоз.
Созревание яйцеклеток и сперматозоидов протекает в основ ном сходным образом, различия возникают только на последней стадии по следующей причине. Для успешного оплодотворения необходимо достаточно большое количество сперматозоидов. Поэтому все четыре образовавшиеся мужские клетки оказыва ются функциональными и жизнеспособными. Основной задачей яйцеклетки является не только оплодотворение, но и успешное созревание плода. С этой целью процесс деления происходит неравноценно: весь желток остается в яйцеклетке, и она ока зывается единственной жизнеспособной. Остальные три клетки вскоре разрушаются посредством апоптоза. Их называют на правительными, или полярными тельцами.
Внешние оболочки яйцеклетки надежно защищают эмбрион, через них, в особенности сквозь скорлупу птичьих яиц, бакте
рии и вирусы не проникают, а |
Подскорлупная |
Зародышевый |
Яичный |
воздух проходит. |
оболочка |
диск (зигота) |
белок |
Сперматозоиды значительно |
Скорлупа |
|
|
меньше яйцеклеток. У млекопи |
|
|
|
тающих они имеют форму длин |
|
|
|
ной нити с головкой, шейкой и |
|
|
|
жгутиком. В головке содержат |
|
|
|
ся хромосомы, а на ее перед |
|
|
|
ней части — аппарат Гольджи |
Белковые |
|
|
|
волокна |
|
|
Строение куриного яйца
90 |
|
|
Глава 4. Размножение организмов |
||
Компекс |
ßäðî |
Головка |
Промежуточный отдел |
Хвост |
|
Гольджи |
|
|
|
|
|
|
|
|
Øåéêà |
|
|
|
|
|
|
|
Осевая нить |
|
|
|
|
|
|
Центриоли Митоондрии
Строение сперматозоида млекопитающих
с ферментами, растворяющими оболочку яйцеклетки для про никновения ядра. Оболочка сперматозоида остается снаружи. Центриоль принимает участие в формировании веретена деления зиготы сперматозоида. Вторая центриоль образует жгутик, по зволяющий сперматозоиду интенсивно передвигаться. Источником энергии для движений жгутика служит как АТФ, синтезируемая митохондриями, так и получаемая в процессе гликолиза. Муж ские гаметы не только вносят генетическую информацию, но и инициируют дальнейшее развитие оплодотворенной яйцеклетки.
1.В чем отличие бесполого размножения от полового? На зовите главное преимущество полового размножения.
2.Перечислите основные типы бесполого размножения.
3.Откуда появляется в дочернем организме пары гомологич ных хромосом при бесполом и половом размножении?
4.Опишите три периода созревания гамет; какой из них на зывают мейозом?
5.Как вы думаете, для чего и почему зародышевый диск в курином яйце всегда оказывается в верхней части желтка?
§ 21. Оплодотворение
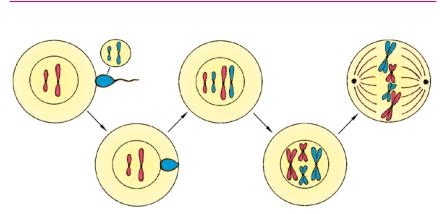
Оплодотворением называют слияние мужской и женской гамет (специализированных половых клеток, имеющих гапло идный набор хромосом), приводящее к образованию оплодот воренной диплоидной яйцеклетки — зиготы. Сущность процесса оплодотворения составляет слияние ядер гамет. Таким образом, при оплодотворении восстанавливается двойной набор, харак терный для соматических клеток. Ядро зиготы содержит две гомологичных хромосомы, то есть любой признак (например, цвет глаз человека или шерстистость собаки) записан в ДНК дважды — в генах отца и в генах матери. После оплодотворения зигота приступает к митотическим делениям — начинается раз витие нового организма.
Оплодотворение, как и гаметогенез, имеет сходные черты у растений и животных.
§ 21. Оплодотворение |
91 |
Оплодотворение у животных. Населяющие планету живые организмы различаются строением, образом жизни, средой оби тания. Одни из них производят очень много половых клеток, другие — относительно мало. Существует разумная закономер ность: чем меньше вероятность встречи мужской и женской гамет и чем меньше выживаемость зиготы и потомства, тем большее число половых клеток продуцируют организмы. Ры бам и земноводным свойственно наружное оплодотворение. Их гаметы попадают в воду, где и происходит оплодотворение. Многие гаметы погибают или поедаются другими существа ми, поэтому эффективность наружного оплодотворения очень низка. Для сохранения вида рыбам и амфибиям необходимо производить огромное количество гамет (треска мечет каждый раз около 10 млн икринок).
Высшие животные и растения используют внутреннее оплодотворение. В этом случае процесс оплодотворения и обра зующаяся зигота защищены организмом матери. Вероятность оплодотворения и выживания потомства значительно повыша ется, поэтому и продуцируется лишь несколько яйцеклеток. Но сперматозоидов все же производится достаточно много, их избыточное количество необходимо для создания вокруг яйце клетки определенной химической среды, без которой оплодот ворение невозможно. Существуют механизмы, препятствующие проникновению лишних сперматозоидов в яйцеклетку. После проникновения одного из них она выделяет вещество, пода вляющее подвижность мужских гамет. Даже если их успевает проникнуть несколько, то с яйцеклеткой сливается только один, остальные гибнут.
Партеногенез (<греч. parthenos девственница). Ряд животных (дафнии, скальные ящерицы, некоторые рыбы, тли) и растения (одуванчик) в определенные периоды способны размножаться
Оплодотворение и начало дробления зиготы у животных
Проникновение |
Удвоение |
Метафаза |
сперматозоида |
хромосом |
первого |
в яйцеклетку |
|
деления |
92 |
Глава 4. Размножение организмов |
без слияния мужской и женской гамет. Развитие происходит из неоплодотворенной яйцеклетки. Диплоидность, например, у скальных ящериц достигается слиянием яйцеклетки с поляр ным тельцем. При этом, как правило, образуются особи только женского пола. Эта разновидность размножения называется партеногенезом. Таким образом, партеногенез — половое, но однополое размножение.
Пчелиная матка откладывает два вида яиц: оплодотворенные диплоидные и неоплодотворенные гаплоидные. Из неоплодотво ренных яиц развиваются трутни, а из оплодотворенных — самки, из которых при хорошем кормлении вырастают матки, а при создаваемом недостатке питания получаются рабочие пчелы.
Иногда партеногенез можно вызвать искусственно, воздей ствуя светом, кислотами, высокой температурой и другими агентами. Если, например, уколоть иглой неоплодотворенную яйцеклетку лягушки, то эта яйцеклетка может начать развитие во взрослую особь. Самопроизвольно партеногенез у лягушек не происходит. Деление яйцеклетки некоторых рыб может на чаться после поверхностного контакта со сперматозоидом рыб близкого вида. Оплодотворения не происходит, но яйцеклетка начинает делиться.
Искусственный партеногенез впервые был осуществлен русским ученым А. А. Тихомировым в 1886 г. посредством об работки яиц тутового шелкопряда серной кислотой, а также механического воздействия. Основным современным способом разведения тутовых шелкопрядов является стимулирование партеногенеза путем кратковременного нагревания яиц до 46 °С. Из неоплодотворенных яйцеклеток развиваются полноценные в генетическом отношении самки шелкопряда. Значительный вклад в изучение генетических механизмов партеногенеза внес ли отечественные ученые академики Б. П. Астауров (1936 г.) и В. А. Струнников (1974 г.).
Оплодотворение у покрытосеменных (цветковых) растений
при общем сходстве с оплодотворением животных имеет некото рые особенности. Процесс формирования мужских гамет носит у них название микроспорогенеза, а женских — мегаспорогенеза. Развитие микроспор происходит в пыльниках. Каждая из че тырех появившихся в результате мейоза микроспор (сравните со сперматогенезом) делится путем митоза и образует два га плоидных ядра: генеративное и вегетативное. Из генеративного ядра в результате второго деления митоза образуется еще два ядра — спермии.
Из четырех мегаспор три дегенерируют (сравните с овоге незом), а четвертая дает начало зародышевому мешку. После периода роста мегаспоры ее гаплоидное ядро обычно делится трижды с образованием восьми гаплоидных ядер. Одно ядро обо

§ 21. Оплодотворение |
93 |
|
Вегетативное ядро |
Прорастающее |
|
Два спермия |
пыльцевое зерно |
|
|
Пыльцевая |
|
|
трубка |
|
Пыльник |
|
|
с пыльцевыми |
Семяпочка |
|
зернами |
||
|
||
Завязь |
Центральное |
|
|
диплоидное |
|
Антиподы |
ÿäðî |
|
|
||
Зародышевый |
Эндосперм |
|
|
||
мешок |
|
|
Синергиды |
Зародыш |
|
|
||
Тычиночная |
Ядро яйцеклетки |
|
íèòü |
|
|
Нектарники |
Двойное оплодотворение |
|
|
||
у покрытосеменных растений |
||
собляется и дает начало ядру яйцеклетки, два других сливают ся, образуя центральное диплоидное ядро. Три ядра (антиподы) перемещаются в дальнюю от пыльцевой трубки часть зародыше вого мешка, еще два ядра (синергиды) располагаются у места проникновения спермия в зародышевый мешок (микропиле).
Пыльцевое зерно, попадая на рыльце пестика, набухает, вегетативное ядро формирует пыльцевую трубку, прорастающую по направлению к семяпочке. Вместе с этой трубкой спермии перемещаются внутрь пестика. Когда кончик трубки касается синергид, трубка разрывается, синергиды разрушаются, а спер мии попадают в зародышевый мешок.
Один из них сливается с гаплоидным ядром яйцеклетки и образует зиготу, из которой формируется зародыш будущего растения. Второй спермий сливается с диплоидным ядром, в результате образуется триплоидное ядро, дающее начало эн досперму. Путем многократных митозов эндосперм формирует питательную среду вокруг зародыша. Таким образом, в одной развившейся многоядерной клетке мегаспоры (зародышевом мешке) происходят два акта оплодотворения.
Второе оплодотворение с образованием и развитием эндоспер ма происходит только после того как оплодотворится яйцеклет ка. Этот универсальный для всех покрытосеменных растений половой процесс носит название двойного оплодотворения. Он открыт в 1898 г. известным русским ботаником С. Г. Навашиным.

94Глава 5. Индивидуальное развитие организмов (онтогенез)
1.В чем заключается генетическая сущность оплодотворения?
2.Как объяснить на молекулярном уровне присутствие у по томства признаков отца и матери?
3.Как происходит оплодотворение у животных?
4.Что называют партеногенезом?
5.Опишите последовательность оплодотворения у растений.
6.Почему оплодотворение покрытосеменных растений на зывают двойным?
Глава 5. ИНДИВИДУАЛЬНОЕ РАЗВИТИЕ ОРГАНИЗМОВ (ОНТОГЕНЕЗ)
Онтогенез (<греч. ontos существо + genesis развитие) — ин дивидуальное развитие особи от зарождения (оплодотворения яйцеклетки, отделения органа вегетативного размножения или деления материнской одноклеточной особи) до конца жизни (смерти или нового деления особи). В ходе онтогенеза проис ходит рост организма, специализация различных групп клеток по функциям, формируется взаимодействие его структур.
Процесс образования целого организма из единственной клетки — зиготы — не перестает удивлять и озадачивать уче ных. Зигота содержит только наследственные задатки — гены, она не обладает характерными признаками и свойствами цело го организма. Каким образом она реализует наследственную информацию и развивается в сложный многофункциональный организм, состоящий из множества разновидных тканей и ор ганов? Выяснение механизма этого развития — одна из самых сложных проблем современной биологии.
Изучением зародышевого развития занимается эмбриология (<греч. embryon зародыш). Еще в IV в. до Р.Х. Гиппократ и Аристотель изучали развитие зародышей животных (в основ ном, кур). Значительные успехи были достигнуты У. Гарвеем («Исследования о зарождении животных», 1651 г.) и К. Ф. Воль фом («Теория зарождения», 1759 г.). Идеи Вольфа были раз виты академиком Петербургской академии наук К. М. Бэром. В 1828 г. он изложил учение об основополагающих структурах эмбриона — зародышевых листках. Карл Бэр доказал, что у всех позвоночных единый план закладки тканей и органов, от четливо проявляющийся на начальных стадиях формирования. Поэтому в период раннего эмбрионального развития все позво ночные очень похожи (закон зародышевого сходства). Признаки класса, рода, вида и, наконец, конкретной особи последователь но проявляются в их строении в процессе дальнейшего роста.
§ 22. Эмбриональное развитие |
95 |
Исследование зародышевых листков продолжил А. О. Ко валевский. Он обнаружил эктодермальные, энтодермальные и мезодермальные листки (слои) у всех групп хордовых. Суще ственный вклад в развитие эмбриологии внес И. И. Мечников.
Начальный период онтогенеза, заканчивающийся выходом организма из яйцевых или зародышевых оболочек, называют эмбриогенезом. Следующий период, продолжающийся до поло вой зрелости, носит название постэмбриогенеза, дорепродуктив ного или ювенильного (<лат. juvenilis юный) периода. Стадия взрослой особи включает репродуктивный (период размножения) и пострепродуктивный периоды. Если на ювенильном этапе продолжается рост и развитие, то на репродуктивной стадии организм представляет собой уже устойчивую систему; пост репродуктивный период завершается старением и смертью.
§ 22. Эмбриональное развитие
На примере представителя хордовых ланцетника рассмотрим основные стадии эмбриогенеза: дробление, гаструляцию, гисто (<греч. histos ткань) и органогенез.
Дробление зиготы. Бластула. Через несколько десятков ми нут после оплодотворения зигота начинает делиться посредством митоза. Сначала возникает вертикальная борозда дробления (перетяжка цитоплазмы), и клетка делится на две одинаковых клетки, называемые бластомерами (<греч. blastos росток + meros доля). Затем другая вертикальная борозда разделяет эти два бла стомера на четыре. Третья борозда проходит горизонтально и син хронно разделяет 4 бластомера на 8. Далее вертикальные и гори зонтальные борозды быстро чередуются, увеличивая количество бластомеров: 16, 32, 64, 128… — до нескольких сотен и тысяч. С ростом количества клеток синхронность дробления теряется.
Дробление происходит очень быстро: у эмбриона лягушки первые тринадцать дроблений занимают всего 6 часов. Быстрота достигается отсутствием роста. Во время короткой интерфазы происходит только репликация ДНК, поэтому процесс и назван дроблением. Бластомеры становятся все мельче и все дальше отходят от центра, образуя полый шарик — бластулу (возник шую полость называют бластоцелью). Начиная с бластулы, клетки зародыша принято называть уже не бластомерами, а эмбриональными клетками. По своим размерам бластула лишь немного больше зиготы, с которой началось дробление. Удвоение ДНК и дробление происходит за счет энергии АТФ и веществ, накопленных заранее в цитоплазме яйцеклетки.
Цитоплазма яйца ланцетника содержит маленький желток и поэтому дробится на приблизительно равные бластомеры.

96 Глава 5. Индивидуальное развитие организмов (онтогенез)
ДРОБЛЕНИЕ |
Полярное тельце |
Бластомеры |
|
|
|
|
|
|
|
Зигота |
|
|
|
Бластоцель |
|
|
|
|
|
|
|
|
|
Бластула |
ГАСТРУЛЯЦИЯ |
|
Эктодерма |
|
Мезодерма |
|
Первичный рот |
|
||
|
|
|
|
|
|
(бластопор) |
|
|
|
|
Первичная |
|
|
|
|
кишка (гастро- |
|
|
|
|
|
öåëü) |
|
|
|
Продольное сечение |
Энтодерма |
Поперечное сечение |
|
|
|
|||
НЕЙРУЛЯЦИЯ |
|
|
|
Хорда |
|
Хордомезодермальный |
Нервная |
||
|
зачаток |
|
трубка |
|
Ранние стадии развития ланцетника
У птиц желток большой, он вытесняет ядро на периферию подобно крупной вакуоли растительной клетки. По этой при чине поперечные борозды дробления зиготы концентрируются на периферийной части яйца, и бластомеры, не содержащие желтка, оказываются существенно мельче. Характерной особен ностью дробления зиготы является отсутствие перемешивания цитоплазмы. Бластомеры оказываются неравноценными, соз давая основу для будущей дифференциации клеток на группы тканей и органов.
Гаструляция — образование двух или трехслойного заро дыша. Когда число бластомеров достигает нескольких сотен или тысяч, у многоклеточных животных наступает следующий
§ 22. Эмбриональное развитие |
97 |
этап эмбриогенеза: образование двухслойного зародыша — га струлы (<греч. gaster желудок). У разных групп животного мира формирование гаструлы происходит с характерными особенностями. Наиболее характерно впячивание одного из по лушарий внутрь другого. Возникающую полость (гастроцель) называют первичным кишечником (его клетки впоследствии образуют пищеварительные органы), а отверстие — первичным ртом (бластопором).
У первичноротых (плоских, круглых и кольчатых червей, моллюсков, членистоногих) бластопор впоследствии становится ротовым отверстием. У вторичноротых (иглокожих, хордовых) ротовое отверстие открывается на противоположном конце за родыша, а бластопор преобразуется в анальное отверстие.
С образованием гаструлы появляются первые признаки диф ференциации клеток. Внешний клеточный слой носит название эктодермы, внутренний — энтодермы (<греч. derma кожа). Клеточные слои эмбриона называют зародышевыми листками. У всех многоклеточных животных, за исключением двухслой ных (губок, кишечнополостных, круглых червей), на границе между эктодермой и энтодермой в процессе гаструляции начи нает обосабливаться и третий зародышевый слой — мезодерма (<греч. mesos средний). Эмбрион становится трехслойным.
Сущность процесса гаструляции состоит в перемещении кле ток и в изменении формы эмбриона. Бластомеры в этот период практически не делятся и не растут.
Гисто и органогенез. Формирование тканей и органов начи нается с нейруляции — формирования отдельных осевых органов: нервной трубки, хорды, кишечной трубки, осевой мезодермы. Продольный фрагмент эктодермы (вдоль будущего позвоночника) вдается внутрь желобком, образуя трубку — зачаток центральной нервной системы. В спинной части зародыша на границе между энтодермой и мезодермой обособляется хордомезодермальный зачаток — будущая хорда.
С самого начала передний конец нервной трубки несколь ко расширен — это будущий головной мозг. По бокам его об разуются два округлых зачатка глаз. Втягивания эктодермы формируют также зачатки органов слуха и обоняния. Из центральной части нервной трубки образуются спинной мозг и периферическая нервная система. Кроме того, из эктодер мы формируются внешние покровные ткани: эпителий кожи, эмаль зубов и пр.
Брюшные энтодермальные клетки образуют трубку — буду щий кишечник. Из выростов этой трубки формируются зачатки печени, легких (плавательный пузырь и внутренние жабры рыб), желез поджелудочной и вилочковой (органа иммунной системы).