

биология 10-11
.pdf68Глава 3. Метаболизм — основа жизнедеятельности клетки
1.Какая группа органических соединений определяет основные свойства организмов? Докажите.
2.Что такое генетический код? Перечислите его свойства.
3.Как происходит транскрипция? Какой принцип лежит в ее основе? Каковы особенности транскрипции у прокариот?
4.В чем состоит функция мРНК?
§ 17. Синтез белковой цепи на рибосоме
Матричные процессы составляют основу способности живых организмов к воспроизведению. В клеточном ядре происходит удвоение ДНК. Новая молекула воспроизводится на матрице старой и представляет собой ее точную копию. Матричная РНК синтезируется на матрице ДНК в виде точной копии одного из участков ДНК (с учетом замены Т → У). Далее на матрице мРНК происходит синтез белков.
Транспортные РНК. Необходимые для синтеза белков ами нокислоты всегда имеются в цитозоле. Они синтезируются или же образуются в процессе расщепления белков в лизосомах. Транспортные РНК присоединяют аминокислоты, доставляют их к рибосоме и осуществляют точную пространственную ори ентацию аминокислот на рибосоме (см. рис. на с. 41).
Рассмотрим устройство тРНК, позволяющее ей выполнять свои сложные функции. В цепочке тРНК, состоящей из 70—90
Антикодон
АБ
ГВ
Ядро |
|
Цитозоль |
|
аминокислоты |
материл |
ìÐÍÊ |
строительный |
|
информация |
|
|
Рибосомы
Аминоислота
Вторичная структура ТРНК |
Схема биосинтеза |
(на примере валиновой) |
белковых молекул |
|
|

§ 17. Синтез белковой цепи на рибосоме |
69 |
Полисома в работе
тРНК
3′-конец
Рибосомы
мРНК
Белок
5′- конец
Аминокислоты
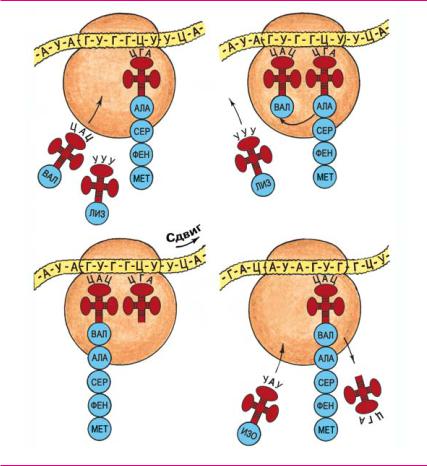
звеньев, имеется 4 пары комплементарных отрезков из 4—7 нуклеотидов — А, Б, В и Г. Комплементарные участки связы ваются водородными связями попарно (как в молекуле ДНК). В результате цепь тРНК «слипается» в четырех местах с об разованием петлистой вторичной структуры, напоминающей лист клевера. В верхушке «листа» располагается триплет (на рис. ЦАЦ), комплементарный тому кодону мРНК, который соответствует транспортируемой аминокислоте (на рис. валин). Если в мРНК код аминокислоты валина ГУГ, то на вершине валиновой тРНК будет триплет ЦАЦ. Комплементарный триплет в тРНК называют антикодоном.
Специальный фермент распознает антикодон тРНК, при соединяет к «черенку листа» определенную аминокислоту (в на шем примере — валин), и затем тРНК перемещает ее к рибосоме. Каждая тРНК транспортирует только свою аминокислоту.
Трансляция (<лат. translatio передача). Перевод последова тельности нуклеотидов мРНК в последовательность амино кислот синтезируемых белков называют трансляцией. Синтез начинается с того, что малая субъединица рибосомы связывается с участком мРНК вблизи 5′ конца, несущим инициирующий кодон АУГ. Далее метиониновая тРНК присоединяется к ко дону АУГ. После этого малая и большая субъединицы, мРНК и инициаторная тРНК объединяются в комплекс, способный синтезировать белок. В процессе синтеза малая субъединица связывает мРНК, а большая катализирует образование пептид ных связей между аминокислотами.
В области кодон антикодоновых взаимодействий рибосомы (активном центре) размещаются два триплета мРНК и соот ветственно две тРНК. Рибосома перемещается по мРНК не плавно, а прерывисто, триплет за триплетом. На каждом шаге присоединяется новая аминокислота. Транспортные РНК пере
70 Глава 3. Метаболизм — основа жизнедеятельности клетки
1 2
мРНК
тРНК
3 |
4 |
Трансляция на этапе присоединения аминокислоты валин
мещаются со своей аминокислотой к рибосоме и «примеряют» свой антикодон к очередному кодону мРНК, находящемуся в активном центре. Между комплементарными нуклеотидами возникают водородные связи. Если антикодон оказывается не комплементарным, то тРНК удаляется в цитоплазму к другим рибосомам. Если же он оказывается комплементарным, то тРНК присоединяется к кодону водородными связями.
Далее синтезируемая белковая цепь присоединяется к при бывшей аминокислоте. Новая тРНК продолжает удерживать всю формирующуюся белковую цепь на кодоне мРНК до прихода следующей тРНК. Освободившаяся тРНК перемещается в цито плазму на поиск своей аминокислоты. Рибосома перемещается на следующий триплет мРНК, и процесс повторяется. Интервал между перемещениями составляет не более 0,2 секунды, а весь синтез белка средней длины продолжается 1—2 минуты.
§ 17. Синтез белковой цепи на рибосоме |
71 |
На одной цепи мРНК могут работать несколько рибосом. На мРНК, содержащей информацию о белке гемоглобине, размеща ется до 5 рибосом, на некоторых других мРНК — до 20 рибосом. Когда в области кодон антикодоновых взаимодействий оказы вается один из трех стоп кодонов УГА, УАА или УАГ, синтез белка завершается. Белковая цепь отделяется от тРНК, а сама рибосома распадается на субъединицы. Рибосомы универсальны
имогут синтезировать белки по любой матрице мРНК. Субъе диницы рибосом соединяются только на время синтеза белка.
Синтез белков требует затрат большого количества энергии. Только на присоединение каждой аминокислоты к тРНК расхо дуется энергия одной молекулы АТФ, а в белке средней длины содержится 400—500 аминокислот. Общее количество затрачи ваемой энергии существенно превышает суммарную химическую энергию образующихся пептидных связей между аминокисло тами. По этой причине белки редко используются клеткой в качестве топлива: слишком велики затраты на их изготовление.
Регуляция транскрипции и трансляции. Клетки различ ных тканей сильно различаются между собой по строению
ифункциям, но все они произошли от единственной зиготы (оплодотворенной яйцеклетки) в результате многократного деле ния. Перед каждым актом деления ДНК реплицируется в две точные копии. Ученым удалось экспериментально доказать, что дифференцированные клетки организма имеют, как правило, одинаковый набор ДНК.
В1945 г. Г. В. Лопашев разработал метод пересадки ядер в яйцеклетку. В 1958 г. Ф. Стюарт впервые вырастил из един ственной клетки взрослой моркови целое растение, а в 1968 г. Дж. Гордону удалось пересадить ядра из клеток кишечника головастика в яйцеклетки лягушки, лишенные собственных ядер, и вырастить из них нормальных лягушек (клонов). Сле довательно, клетки всех тканей организма имеют один и тот же набор генов, одну и ту же информацию о строении белков.
Однако клетки каждой ткани производят свои белки, свои ферменты. Белковый гормон роста синтезируется только в клет ках гипофиза, зрительный белок опсин — в клетках сетчатки глаза, а инсулин — в клетках поджелудочной железы.
Такое разнообразие существует потому, что клетки каждой ткани реализуют только свою часть информации ДНК. В клет ках разных тканей происходит транскрипция разных генов, син тезируются разные мРНК, по которым воспроизводятся разные белки. Набор активно работающих генов зависит не только от тканевой принадлежности клетки, но и от периода ее жизненно го цикла. Так, за интенсивное деление и рост клеток в период развития эмбриона отвечают определенные гены, в большин стве клеток взрослых существ эти гены молчат (их включение

72 |
Глава 3. Метаболизм — основа жизнедеятельности клетки |
может привести к появлению опухолей). В эукариотической клетке многоклеточного организма функционирует около 5 % имеющихся у нее генов, остальные находятся в репрессированом состоянии (не активны). Эти факты свидетельствуют о том, что активность генов регулируется.
Основным механизмом регуляции активности генов (экс прессии) служит затруднение или стимуляция присоединения РНК полимеразы к промотору для начала транскрипции. Раз личают два вида регуляторных белков.
Белки репрессоры, связываясь с оператором (регулирующий участок ДНК, расположенный в промоторе или между ним и геном), препятствуют присоединению полимеразы к промотору. Белки активаторы, напротив, облегчают присоединение поли меразы и стимулируют таким образом транскрипцию. Деятель ностью регуляторных белков управляют вещества эффекторы. Связываясь с белками регуляторами, эффекторы изменяют их биологическую активность, «включая» и «выключая» транс крипцию. Эффекторами часто служат гормоны. Самые активные промоторы у бактериофагов, поэтому они и побеждают генети ческий механизм клетки хозяина, заставляя ее синтезировать преимущественно свои белки.
У прокариот гены, кодирующие синтез ферментов, участвую щих в одной цепи биохимических превращений (например, осуществляющих расщепление глюкозы), зачастую собраны под одним промотором (с него начинается транскрипция). Такую группу структурных генов (определяющих структуру ферментов) вместе с промотором, оператором, последовательностями начала
иконца транскрипции называют опероном. По мере необходи мости опероны «включаются» и «выключаются». Так, кишечная палочка использует для жизнедеятельности и глюкозу и лактозу (молочный сахар), но если она находится в среде, содержащей только глюкозу, то не растрачивает энергию АТФ на синтез бел ков, расщепляющих лактозу. Ген регулятор лактозного оперона синтезирует белок репрессор, который, взаимодействуя с операто ром, препятствует присоединению РНК полимеразы к промотору
и«выключает» транскрипцию генов, в которых закодированы ферменты, необходимые для усвоения лактозы. Если же бакте рии переместить в среду, содержащую только лактозу, то лакто за, попадая в клетки, инактивирует белок репрессор и начина ется транскрипция оперона, отвечающего за усвоение лактозы.
Генетический аппарат эукариот устроен намного сложнее, чем у прокариот, именно вследствие большей сложности ре
Оперон — участок ДНК у прокариот для транскрипции на одну мРНК
Промотор Оператор |
ГЕН 1 |
ГЕН 2 |
ГЕН 3 |
Терминатор |

§ 17. Синтез белковой цепи на рибосоме |
73 |
гуляции генетических процессов. У человека генов в 2 раза больше, чем у дрозофилы, и в 5 раз больше, чем у дрожжей. У высших животных и человека не более 5 % ДНК кодиру ют собственно гены (у прокариот — более 50 %). Значительно больше места в ДНК занимают разнообразные регуляторные элементы. Основную же часть генома (набора генов организма) эукариот составляет молчащая ДНК — многократно повторяю щиеся короткие последовательности нуклеотидов, не кодирую щие макромолекулы.
Матричные РНК большинства генов эукариот кроме участ ков, непосредственно кодирующих белки (экзонов), содержат фрагменты, подлежащие удалению (интроны). Специальные ферменты в ядре вырезают интроны и сшивают между собой экзоны. Этот процесс называют сплайсингом (<англ. splice сра щивать канаты без узлов, морской термин). После сплайсинга и некоторых других операций мРНК называют зрелой, далее она поступает в цитоплазму к рибосомам.
Ферменты не всегда сшивают экзоны в том порядке, в каком они были считаны с ДНК. Измененный порядок сшивки экзонов называют альтернативным сплайсингом. Во многих случаях альтернативный сплайсинг имеет тканеспецифичный характер:
всозревших мРНК разных тканей и органов порядок экзонов может несколько различаться, соответственно отличаются и транслируемые белки. У человека и высших животных около 40 % генов подвергаются альтернативному сплайсингу. Так эко номно устроена ДНК: ген один, а белков получается несколько. Функции этих белков обычно очень сходны, поэтому их на зывают изоформами, но в ряде случаев такие белки ферменты обеспечивают разные процессы метаболизма.
Урастений и многоклеточных животных некоторые интроны содержат гены особых микроРНК (миРНК), включающие всего лишь 20—28 нуклеотидов. Эти миРНК играют важную роль
врегуляции сотен генов. Нуклеотидная последовательность миРНК комплементарна участку мРНК, трансляцию которой она регулирует. Связываясь с таким участком, миРНК либо подавляет трансляцию, либо вызывает разрушение мРНК.
Подготовка кодирующего участка мРНК к трансляции у эукариот
|
|
мРНК после транскрипции |
||
Экзон 1 |
Интрон 1 |
Экзон 2 |
Интрон 2 |
Экзон 3 Интрон 3 Экзон 4 |
→Подготовка мРНК к трансляции: вырезание интронов и сшивка (сплайсинг) экзонов |
||||
Экзон 1 |
Экзон 2 |
Экзон 3 |
Экзон 4 Зрелая мРНК (готовая к трансляции) |
|
Варианты альтернативного сплайсинга (наиболее распространено отсутствие какого-то экзона) |
|||||
|
|
|
→ |
→ |
|
|
|
|
|
|
|
Экзон 1 |
Экзон 2 |
Экзон 4 |
|
Экзон 1 |
Экзон 3 Экзон 4 |

74 |
Глава 3. Метаболизм — основа жизнедеятельности клетки |
Существуют системы, регулирующие синтез веществ в орга низме как едином целом. В клетках желез внутренней секреции вырабатываются соответствующие гормоны, которые разносятся с кровью по всему телу. Они регулируют процессы синтеза мРНК и трансляцию именно в тех клетках, для которых они предназначены. На поверхности клеток есть рецепторы для «сво их» гормонов. Связываясь с рецепторами, гормоны управляют активностью различных систем клетки, регулирующих обмен. В результате может включаться выключаться как транскрипция конкретных генов, так и синтез белков на рибосомах.
Даже синтезированные мРНК могут долгое время не транс лироваться, если нет команды от гормонов. Каждый гормон че рез определенные системы клеток активирует свои гены. Уско рение синтеза необходимого белка достигается более активным присоединением рибосом к мРНК — созданием полисомного ком плекса (нескольких рибосом, перемещающихся друг за другом).
Как транскрипция, так и трансляция могут подавляться различными химическими веществами, относящимися к классу антибиотиков (<греч. anti против + bios жизнь). Например, эри тромицин и стрептомицин подавляют синтез белка на рибосомах бактерий. Бледная поганка содержит вещество, подавляющее функционирование РНК полимеразы человека. Прекращение полимеразой синтеза мРНК приводит к тяжелейшим послед ствиям.
Подведем итоги. В воспроизведении белков участвуют: ДНК; мРНК; тРНК для 20 аминокислот; рибосомы, состоящие из нескольких рРНК и десятков различных белков; целый ком плекс ферментов. Для осуществления синтеза белковой моле кулы средней длины необходим тонкий, специфический подвод энергии посредством около 1 000 молекул АТФ. Практически вся клетка участвует в синтезе, нарушение строения хотя бы одного из компонентов нарушает процесс воспроизведения бел ковых молекул.
Для современных ученых удивителен сам факт функциони рования этой сложной системы в организме. Возможность же ее самопроизвольного появления многие исследователи абсолютно исключают. Познание внутриклеточных процессов приводит к мысли о Создателе.
1.Каково предназначение матричных процессов в клетке?
2.Опишите строение и функции тРНК.
3.Опишите последовательность трансляции белков.
4.Способна ли каждая рибосома синтезировать любые белки? Может ли каждая тРНК переносить любые аминокислоты?
5.Почему необходимо «включать» и «выключать» гены? Ка ковы механизмы этих процессов?


76 |
Глава 4. Размножение организмов |
Глава 4. РАЗМНОЖЕНИЕ ОРГАНИЗМОВ
Наш повседневный опыт печально свидетельствует о том, что все живое подвержено смерти.* Существа болеют, старятся
инаконец умирают. У многих жизнь еще более короткая: их съедают хищники. Чтобы жизнь на Земле не прекратилась, все существа наделены универсальным свойством — способностью к размножению.
При всем разнообразии живых организмов, населяющих пла нету, при всех различиях в строении и образе жизни, способы их размножения в природе сводятся к двум формам: бесполой
иполовой. Некоторые растения сочетают эти две формы, раз множаясь бесполым путем (клубнями, черенками, отводками и т. д.) и одновременно половым (семенами).
Вслучае бесполого размножения потомство развивается из клеток исходного организма. При половом размножении развитие нового существа начинается с единственной клетки, образовавшейся от слияния двух родительских — мужской и женской.
Сущность размножения состоит в сохранении не только жизни в целом, но и каждого отдельного вида животных и растений, в организации преемственности между потомством
иродительскими существами. Молекулярную основу процессов размножения всех организмов составляет способность ДНК к самоудвоению. В результате генетический материал воспроиз водится, а строение и функционирование дочерних организмов оказывается таким же, что и родительских.
* Священное Писание и творения святых отцов пронизаны мыслью о том, что смерть и тление не были сотворены изна чально, а вошли в мир вследствие грехопадения первого человека (Прем. 1,13 и 2,23, Рим. 5,12, приложения с. 344, и т. д.).
§ 18. Деление соматических клеток. Митоз |
77 |
§ 18. Деление соматических клеток. Митоз
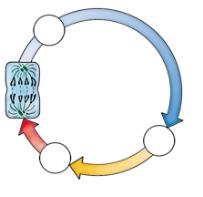
Клеточный цикл. Процесс деления и интерфаза (промежу ток между делениями) тесно взаимосвязаны, их совокупность составляет клеточный цикл. Его продолжительность в клет ках растений и животных составляет в среднем 10—20 часов. В химически активной среде пищевого тракта клетки эпителия кишечника быстро изнашиваются и поэтому делятся дважды в сутки, клетки роговицы глаза приступают к делению один раз в трое суток, а клетки эпителия кожи — раз в месяц. Нейроны и некоторые лейкоциты никогда не делятся. В печени живот ных находятся покоящиеся клетки, которые делятся только, например, при удалении части печени.
После разделения клетки на две дочерних в каждой из них начинается первый, самый продолжительный (от 10 часов до нескольких суток), период клеточного цикла — пресинтетический (G1). Клетка готовится к удвоению хромосом и растет: синтези руются белки, РНК, увеличивается количество рибосом и мито хондрий, поверхность ЭПС. В течение следующих 6—10 часов синтетического периода (S) происходит синтез ДНК (репликация). Третий постсинтетический период (G2) занимает 3—6 часов, клетка готовится к делению: синтезируются белки микротрубо чек, запасаются вещества. В интерфазе, состоящей из периодов G1, S, G2, происходят все основные процессы обмена веществ.
Четвертый, последний период клеточного цикла, митоз, длится в среднем от 1 до 3 часов в зависимости от внешних условий (освещения, температуры и пр.).
Деление клеток происходит в два этапа. Сначала разделя ются хромосомы, а затем происходит цитокинез — разделение цитоплазмы.
Митоз. Основной способ деления эукариотических клеток называют митозом (<греч. mitos нить). Различают четыре фазы митоза: профаза, метафаза, анафаза и телофаза.
Профаза (<греч. pro перед). В профазе заканчивается при
готовление к делению. Хромосомы |
|
|
|
||
утолщаются и становятся видимы |
2n2C |
|
|
||
в световой микроскоп. К концу |
|
G1 |
|||
|
|
||||
профазы они представляют собой |
|
|
|||
плотно упакованные дочерние хро |
|
|
|
||
матиды, образовавшиеся в процессе |
Mèòîç |
|
|
||
удвоения ДНК и соединенные цен |
|
|
|||
|
|
|
|||
тромерами. |
G2 |
|
|
||
|
|
S |
2n2C |
||
Периоды клеточного цикла. |
|||||
|
|||||
4n4C |
|
||||
2n, 4n — набор хромосом, |
|
|
|||
2С, 4С — число молекул ДНК в наборе