

24.Изометрические и изотонические сокращения
Изометрические-при неизменной длине мішці(при попітке поднять непосильній груз).Условие:постоянная длина при максимальном тонусе
Изотонические-сокращение мішці без нагрузки.Изменяется длина при неизменном тонусе
не при изометрическом,ни при изотоническом не будет віполнятся работа.поєтому скелетніе мішці в нормальніх условиях сокращаются смешано
25.Утомление мішці
Несколько теорий утомления на уровне изолированой мішці:
1)теория истощения-утомление обьясняется у меньшением в мішце питательніх веществ,АТФ,креатинин,фосфат,гликоген и т.д
2)теория засорения-обьясняет утомление накоплением метаболитов(молочная кослота,СО2,К+,продукті обмена азота)
3)теория удушения-снижение кислорода,что тормозит ресинтез АТФ
Все теории правильніе,но в зависимости от условий тот или иной фактор начинает преобладать
первім утомлении наступает в синапсе(из-за нарушения синтеза медиатора при длительном раздражении и повішеном расходовании)
В условиях целого организма утомление первім наступает в корковіх двигательніх ентрах.связано с большим количеством синапсов и с процессами торможения в нервніх клетках.
26.Синапсі цнс
Синапсі ЦНС- єто специализированіе межклеточніе контакті между нейронами
По взаимоотношению между различніми отделами нейронов деляться на:
-аксо-дендритические
-аксо-соматические
-аксо-аксональніе
-дендро-дендритические
По механизму передачи возбуждения:
1)синапсі с химическим механизмом передачи.имеют синаптическую щель,пузірьки медиаторов в пресинаптической части,в постсинаптической-холинорецепторі
2)синапсі с єлектрическим механизмом передачи-у человека только в ємбриогинезе.нет пузірьков с медиатором,синаптической щели,хеморецепторов.большой размер
3)синапсі со смешанім типом передачи
Особенности синапсов в цнс:нейромодуляция-регуляция віделения основного медиатора с помощью нейропептидов и нейрогормонов(с пре- и постсинаптическим действием
27.
Нейромедиаторы – это вещества, образующиеся в пресинаптических нервных окончаниях, хранящиеся там в особых везикулах, выделяющиеся из нервных окончаний под действием нервного импульса в синапс, связывающиеся со специфическим рецептором на постсинаптической мембране и имеющие механизмы для быстрого удаления медиаторов из синаптической щели. В роли медиаторов выступают ацетилхолин, дофамин, норадреналин, серотонин, ГАМК, глицин и некоторые другие вещества.В зависимости от эффектов на постсинаптической мембране нейромедиаторы делят на возбуждающие и тормозные.К возбуждающим нейромедиаторам относят ацетилхолин, дофамин, серотонин, глутаминовую кислоту, норадреналин. В некоторых синапсах в качестве возбуждающего нейромедиатора могут выделяться пурины, АТФ, некоторые нейропептиды.К тормозным нейромедиаторам относят гамма-аминомасляную кислоту (ГАМК) и глицин. Критерии нейромедиаторов:
1) вещество должно синтезироваться и накапливаться в пресинаптических нервных окончаниях, из которых должно выделяться в ответ на деполяризацию;
2) выделившийся нейромедиатор должен оказывать постсинаптическое действие путем взаимодействия со специфическим постсинаптическим рецептором;
3) вещество должно или разрушаться в синаптической щели, или удаляться из нее с помощью механизма обратного захвата;
4) введение этого вещества в нервную ткань должно оказывать такое же действие, как и эндогенное вещество.
Кроме медиаторов, в синапсе могут выделяться также нейромодуляторы. Нейромодуляторы – это вещества, которую прямо не изменяют состояние синаптических мембран, но влияют на интенсивность и продолжительность действия классических нейромедиаторов и таким образом влияют на активность синаптической передачи сигнала. Чаще всего в роли нейромодуляторов выступают нейропептиды. Нейропептиды – это обширная группа короткоцепочечных пептидов, которые могут выступать и как нейромедиаторы, и как нейромодуляторы.
Например, энкефалины и эндорфины связываются со специфическими рецепторами (с которыми также взаимодействует морфин), при этом происходит подавление чувства боли. Другой пептид – вещество Р, подавляет ощущение боли, а также вызывает сокращение гладких мышц. Ангиотензин II – гормон местного действия, сильно влияет на кровеносные сосуды и работу ЦНС. Так же действует и вазоактивный кишечный пептид (ВИП).
Простагландины действуют как локальные химические агенты, так как они очень быстро инактивируются. Модулируя синаптическую передачу, они влияют на многие физиологические процессы, например, меняют секрецию медиаторов, работу аденилатциклаз.
28.
При приходе электрического нервного импульса в пресинаптическое нервное окончание происходит деполяризация пресинаптической мембраны, в ней открываются потенциалзависимые Са-чувствительные каналы, и из синаптической щели в пресинаптическое окончание поступают ионы Са2+, необходимые для активации везикул. Последние прилипают к пресинаптической мембране, и из них путем экзоцитоза в синаптическую щель порциями (квантами) выходит медиатор. Квант медиатора диффундирует к постсинаптической мембране и взаимодействует со специфическим рецептором, изменяя его конформацию, вследствие чего открываются натриевые каналы. Na+ входит внутрь мышечной клетки, вызывая деполяризацию, а К+ по градиенту начинает выходить наружу, вынося излишек положительного заряда. Так возникает возбуждающий постсинаптический потенциал (ВПСП), или потенциал концевой пластинки (ПКП), по механизму являющийся локальным ответом. Эти потенциалы могут суммироваться по количеству и по времени. Когда ВПСП достигает критического уровня, в соседних участках мембраны за счет локального кругового электрического тока активируются потенциалзависимые натриевые каналы, что и приводит к развитию потенциала действия (ПД). Он не может возникнуть в самой постсинаптической мембране, так как в ней нет потенциалзависимых каналов. Таким образом осуществляется передача сигнала с помощью возбуждающих нейромедиаторов.
29.
Свойства нервных центров:
Одностороннее проведение возбуждения: обусловлено особенностями расположения и характером функционирования синапсов в нервных центрах.
Замедление (задержка) распространения возбуждения: обусловлено наличием множества синапсов на пути следования сигнала – может составлять от 0,5 до 2-4 мс в зависимости от количества синапсов, участвующих в работе данного участка нервной цепи.
Иррадиация возбуждения – распространение возбуждения от активной клетки на соседние клетки и центры – обусловлено наличием множества ветвлений аксонов.
Суммация возбуждений (или торможений): временная и пространственная суммация сигналов, каждый из которых может быть ниже пороговых значений, но в результате суммации может привести к возникновению ПД. Временная суммация обусловлена тем, что ВПСП от предыдущего импульса еще продолжается, когда приходит следующий импульс. Многие нейронные процессы имеют ритмический характер и таким образом могут суммироваться, давая начало надпорогому возбуждению в нейронных цепях. Пространственная суммация заключается в том, что раздельная стимуляция каждого из двух аксонов вызывает образование подпорогового ВПСП, тогда как при при одновременной стимуляции обоих аксонов возникает ПД, обусловленный конвергенцией их влияний. Чаще всего наблюдается пространственно-временная суммация, когда на центральном нейроне сходятся импульсы, поступающие от разных нейронов в достаточно близкие микроинтервалы времени, в результате чего их воздействия суммируются.
Тонус нервных центров – наличие постоянного уровня возбуждения в части нейронов нервного центра. Тонус (фоновая активность) объясняется следующими причинами: а) спонтанной активностью нейронов ЦНС; б) гуморальными влияниями биологически активных веществ, циркулирующих в крови и влияющих на активность нейронов; в) импульсацией от различных рефлексогенных зон; г) суммацией миниатюрных потенциалов, возникающих в результате выделения квантов медиатора из аксона; д) циркуляцией возбуждения в ЦНС.
Трансформация ритма возбуждения – это изменение числа импульсов, возникающих в нейронах центра на выходе относительно числа импульсов, поступающих на вход данного центра. Уменьшение может быть вследствие уменьшения возбудимости за счет процессов пре- и постсинаптического торможения. При большом потоке афферентных влияний, когда уже все нейроны нервного центра возбуждены, дальнейшее увеличение афферентных входов не приводит к увеличению числа возбужденных нейронов.
Последействие (например, длительное циркулирование импульсов по «нейронной ловушке»), или «облегчение» проведения. Следовые процессы имеют важное значение в понимании механизмов памяти. Непродолжительное последействие до 1 часа связано с циркуляцией импульсов в нейронных цепях и обеспечивает краткосрочную память. Следовые процессы в спинном мозге длятся несколько секунд или минут, в подкорковых центрах – десятки минут, часы, дни, в КБП – до нескольких десятков лет.
Утомляемость нервных центров – наблюдается при частых повторных раздражениях и связана с истощением запасов нейромедиаторов в пресинаптических окончаниях, снижением чувствительности рецепторов.
Пластичность (приспособляемость, смещение функций) – способность нервного центра к функциональным перестройкам. Например, способность осуществлять функцию при повреждении части нейронов за счет того, что оставшиеся нейроны замещают (компенсируют) функцию погибших. Другим проявлением пластичности является синаптическое облегчение – улучшение проведения в синапсах после короткого раздражения афферентных путей. Облегчение достигает максимума, когда импульсы поступают с интервалом в несколько миллисекунд. Главная причина этого облегчения, по-видимому, заключается в накоплении ионов кальция в пресинаптическом нервном окончании, когда ионный насос не успевает его выводить. Соответственно увеличивается высвобождение медиаторов, ускоряется их синтез, увеличивается активность рецепторов. Следующим проявлением пластичности является образование временных связей, которые обеспечивают образование условных рефлексов.Конвергенция или принцип общего конечного пути – схождение, поступление к одному нейрону импульсов от различных нервных центров и разных рецепторов («воронка» Шеррингтона). На одном нейроне ЦНС может быть до 20 тысяч синапсов (например, на мотонейроне спинного мозга). Конвергенция может облегчать процесс возникновения возбуждения на общем нейроне в результате пространственной суммации подпороговых ВПСП либо блокировать его вследствие преобладания тормозных влияний. Благодаря конвергенции один мотонейрон может участвовать в нескольких различных реакциях, разные раздражители могут вызвать однотипную реакцию (например, поворот головы, настораживание может быть в ответ на действие, световых, звуковых и других раздражителей; рефлексы чихания, сосания, глотания, кашля заканчиваются на одних и тех же мотонейронах, иннервирующих мышцы глотки).Дивергенция (иррадиация) – способность нервной клетки устанавливать многочисленные синаптические связи с различными нервными клетками, что объясняется ветвлением аксонов нейронов (в среднем нейрон образует до 1000 окончаний) и их способностью устанавливать многочисленные связи с другими нейронами, наличием вставочных нейронов, аксоны которых тоже ветвятся. Благодаря дивергенции возможна иррадиация (распространение) возбуждения и быстрое вовлечение в ответную реакцию многих центров, расположенных на разных уровнях ЦНС. Дивергенция обеспечивает иррадиацию возбуждения в центральных нервных образованиях. Тормозные процессы ограничивают дивергенцию и делают процессы управления более точными. Когда торможение снимается, то может произойти резкое преобладание процессов возбуждения и нарушение деятельности ЦНС (например, при столбняке).


30.
Важнейший принцип, на котором основана работа нервной системы, – это принцип рефлекторной регуляции. Впервые представление о рефлекторном характере деятельности головного мозга было разработано Сеченовым, развито И.П.Павловым, В.М.Бехтеревым, Ч.Шеррингтоном, П.К.Анохиным. Они показали, что в нижележащих отделах ЦНС – подкорковых ядрах, стволе, спинном мозге – рефлекторные реакции осуществляются по прирожденным, наследственно закрепленным нервным путям, а в КБП нервные связи вырабатываются, создаются в процессе индивидуальной жизни животных и человека, в результате сочетания многочисленных действующих на организм раздражителей.
Рефлекс – это стереотипная ответная реакция организма на раздражение, осуществляемая при участии ЦНС. Рефлекторная дуга – это совокупность структур, при помощи которых осуществляется рефлекс (рисунок 2). Чаще всего она состоит из пяти звеньев:
1) периферические рецепторы, к которым подходят окончания афферентного (чувствительного) нейрона;
2) афферентный (чувствительный, центростремительный) нейрон – воспринимает изменения внешней или внутренней среды организма. Совокупность рецепторов, раздражение которых вызывает рефлекс, называется рефлексогенной зоной;
3) вставочный (ассоциативный) нейрон, расположенный в спинном или головном мозге – обеспечивает связь с другими отделами ЦНС, переработку и передачу импульсов к эфферентному нейрону;
4) эфферентный (двигательный, центробежный) нейрон – вместе с другими нейронами перерабатывает информацию, сформировывает ответ в виде нервных импульсов;
5) эффектор (исполнитель) – рабочий орган.
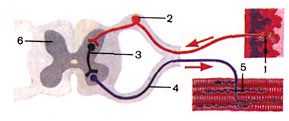
Рисунок 2 – Схема рефлекторной дуги:
1 – рецептор; 2 – центростремительный нейрон; 3 – вставочный нейрон;
4 – центробежный нейрон; 5 – нервные окончания в мышце; 6 – спинной мозг.
Большинство рефлексов замыкается в головном и спинном мозге, и лишь небольшое количество их замыкается вне ЦНС – в вегетативных ганглиях. Вставочных нейронов может быть от одного до множества (в нервных центрах).
Самая простая рефлекторная дуга – моносинаптическая. Она состоит из двух нейронов – афферентного и эфферентного. Таких рефлексов мало – как правило, это сухожильные рефлексы (например, спинальные миостатические – возникающие в ответ на растяжение мышцы). Чаще дуга рефлекса содержит не менее трех нейронов: афферентный, вставочный и эфферентный. Подобные дуги называют полисинаптическими.
Классификация рефлексов:
1. По способу образования:
1) безусловные – врожденные;
2) условные – приобретенные.
2. По компонентам синаптической дуги:
1) моносинаптические;
2) полисинаптические.
3. По уровню замыкания рефлекса:
1) спинальные;
2) бульбарные;
3) мезенцефальные;
4) таламические;
5) кортикальные и т. д.
4. По характеру рецепторов:
1) интероцептивные;
2) экстероцептивные;
3) проприоцептивные.
5. По биологическому значению:
1) половые;
2) оборонительные;
3) пищевые и др.
6. По участию соматических или вегетативных отделов ЦНС:
1) соматические;
2) вегетативные.
7. По конечному результату:
1) сердечные;
2) сосудистые;
3) слюноотделительные и т. д.
31.
Тормозные нейроны — тип интернейронов, аксоны которых образуют на телах и дендритах возбуждающих клеток нервные окончания, в которых вырабатывается особый тормозный медиатор. Нервные импульсы, возникающие при возбуждении Т. н. не отличаются от потенциалов действия обычных возбуждающих нейронов. Однако в нервных окончаниях Т. н. под влиянием приходящего по аксону импульса выделяется медиатор, который не деполяризует, а, напротив, гиперполяризует постсинаптическую мембрану. Гиперполяризация регистрируется в форме электроположительной волны, обозначаемой как тормозный постсинаптический потенциал. Примером тормозных нейронов являются клетки Реншоу в спинном мозге, грушевидные нейроны (клетки Пуркинье) коры мозжечка, звездчатые клетки коры мозга и др.
ТПСП возникает при действии в синапсах глицина, гамма-аминомасляной кислоты. ТПСП может развиваться и под действием медиаторов, вызывающих ВПСП, но в этих случаях медиатор вызывает переход постсинаптической мембраны в состояние гиперполяризации.
Его появление связывают с выделением в синаптическую щель специфического медиатора. В синапсах разных нервных структур роль тормозного медиатора могут выполнять различные вещества. В ганглиях моллюсков роль тормозного медиатора выполняет ацетилхолин, в ЦНС высших животных — гамма-аминомасляная кислота, глицин.

32.
Классификация торможения:
I. По локализации места приложения в синапсе:
1 – пресинаптическое торможение – наблюдается в аксо-аксональных синапсах, блокируя распространение возбуждения по аксону (в структурах мозгового ствола, в спинном мозге). В области контакта выделяется тормозной медиатор (ГАМК), вызывающий гиперполяризацию, что нарушает проведение волны возбуждения через этот участок.
2 – постсинаптическое торможение – основной вид торможения, развивается на постсинаптической мембране аксосоматических и аксодендрических синапсов под влиянием выделившихся ГАМК или глицина. Действие медиатора вызывает в постсинаптической мембране эффект гиперполяризации в виде ТПСП, что приводит к урежению или полному прекращению генерации ПД.
II. По эффектам в нейронных цепях и рефлекторных дугах:
1 – реципрокное торможение – осуществляется для координации активности мышц, противоположных по функции (Шеррингтон). Например, сигнал от мышечного веретена поступает с афферентного нейрона в спинной мозг, где переключается на α-мотонейрон сгибателя и одновременно на тормозной нейрон, который тормозит активность α-мотонейрона разгибателя.
2 – возвратное торможение – осуществляется для ограничения излишнего вобуждения нейрона. Например, α-мотонейрон посылает аксон к соответствующим мышечным волокнам. По пути от аксона отходит коллатераль, которая возвращается в ЦНС – она заканчивается на тормозном нейроне (клетка Реншоу) и активирует ее. Тормозной нейрон вызывает торможение α-мотонейрона, который запустил всю эту цепочку, то есть α-мотонейрон сам себя тормозит через систему тормозного нейрона.
3 – латеральное торможение (вариант возвратного). Пример: фоторецептор активирует биполярную клетку и одновременно рядом расположенный тормозной нейрон, блокирующий проведение возбуждения от соседнего фоторецептора к ганглиозной клетке («вытормаживание информации».
33.
Доминанта – это очаг возбуждения, наиболее важного для организма в данный момент и подчиняющего себе (притягивающего к себе импульсацию) активность других нервных центров, преобладание активности центров рефлекса, который в данный момент наиболее важный, и затормаживание в этот момент других. Доминирующий очаг может быть в состоянии возбуждения долго, без признаков утомления. Это осуществляется за счет следовых процессов, обусловленных суммацией ВПСП, вызванных приходящими к нейронам подпороговыми нервными импульсами; синаптической потенциацией (облегчением) при ритмическом раздражении пресинаптических входов; циклическими связями, способными обеспечить следовую стимуляцию нервных центров. Доминантное состояние обеспечивается суммацией значимых раздражителей и сопряженным торможением конкурирующих центров. Доминантный очаг возбуждения обладает такими свойствами, как инерционность, стойкость, повышенная возбудимость, способность «притягивать» к себе иррадиирующие возбуждения и оказывать угнетающие влияния на центры-конкуренты. Значение доминантного очага возбуждения в ЦНС заключается в том, что на его базе формируется конкретная приспособительная деятельность, ориентированная на достижение полезных результатов. Именно благодаря доминанте возможно сосредоточение психической деятельности (внимание) и выполнение умственной или физической деятельности. В период поиска пищи и поедания возникает пищевая доминанта. Существуют половая, оборонительная и другие виды доминант.
Принцип обратной связи (П.К.Анохин) – замыкание рефлекторной дуги в рефлекторное кольцо или рефлекторную цепь в результате того, что полученный в ходе осуществления рефлекса результат сравнивается с результатом, оптимальным для организма в данный момент, и происходит коррекция рефлекторной реакции для получения оптимального эффекта. Это основной механизм, обеспечивающий координацию ЦНС и приспособление организма к изменяющимся условиям внешней и внутренней среды. С помощью обратной связи осуществляется саморегуляция функций в организме, например, поддержание артериального давления, регуляция дыхания за счет хеморецепторов дыхательного центра (реагируют на содержание СО2).
34.
Двигательные системы спинного мезга,
их организация, схема.
вигательные
системы спинного мозга обеспечивают
реализацию влияний центров головного
мозга в управлении опорно-двигательным
аппаратом, а также осуществляют
собственные рефлексы и регуляцию
мышечного тонуса туловища, шеи и
конечностей. Если в качестве объединяющего
стержня рефлексов конечностей принять
характер ответной реакции, то все их
можно объединить в четыре группы: 1)
сгибательные, 2) разгибательные, 3)
ритмические и 4) рефлексы позы.
35. Фазический рефлекс на растяжение
Афферентный аксон группы Iаотмышечного веретенапрямой мышцы бедра входит вспинной мозги разветвляется. Его ветви поступают всерое вещество спинного мозга. Некоторые из них оканчиваются непосредственно (моносинаптически) наальфа-мотонейронах, посылающих двигательные аксоны к прямой мышце бедра (и к ее синергистам, таким как промежуточная широкая мышца бедра), разгибающей ногу в колене.Аксоныгруппы Iа обеспечивают моносинаптическое возбуждение альфа- мотонейрона. При достаточном уровне возбуждения мотонейрон генерирует разряд, вызывающий сокращение мышцы. Другие ветви аксона группы Iа образуют окончания на тормозных интернейронах той же группы. Эти тормозные интернейроны оканчиваются на альфа- мотонейронах, иннервирующих мышцы, которые соединены с подколенным сухожилием(в том числе полусухожильную мышцу), - антагонистические мышцы-сгибатели колена. При возбуждении тормозных интернейронов Iа подавляется активность мотонейронов мышц-антагонистов. Таким образом, разряд (стимулирующая активность) афферентов группы Iа от мышечных веретен прямой мышцы бедра вызывает быстрое сокращение этой же мышцы и сопряженное расслабление мышц, соединенных с подколенным сухожилием.Рефлекторная дугаорганизована так, что обеспечивается активация определенной группыальфа-мотонейронови одновременное торможение антагонистической группы нейронов. Это называетсяреципрокной иннервацией. Она свойственна многим рефлексам, но не единственная для систем регуляции движений. В некоторых случаях двигательные команды вызывают сопряженное сокращение синергистов и антагонистов. Например, при сжимании кисти в кулак мышцы-разгибатели и мышцы-сгибатели кисти сокращаются, фиксируя положение кисти.
Импульсный разряд афферентов группы Iа наблюдается, когда врач наносит неврологическим молоточком легкий удар по сухожилию мышцы, обычно четырехглавой мышцы бедра. Нормальная реакция - кратковременное мышечное сокращение. При патологических изменениях возбудимости альфа- мотонейронов фазический рефлекс растяжения оказывается подавленным либо усиленным. Раньше этот рефлекс называли глубоким сухожильным рефлексом. Однако такой термин ошибочен, поскольку за рефлекс ответственны мышечные рецепторы, а не сухожильные. Сухожилие лишь обеспечивает быстрое растяжение мышцы.
36.
Фазные рефлексы — это однократное сгибание конечности при однократном раздражении кожи или проприорецепторов. Одновременно с возбуждением мотонейронов мышц-сгибателей происходит реципрокное торможение мотонейронов мышц-разгибателей. Рефлексы, возникающие с рецепторов кожи, являются по-лисинаптическими, они имеют защитное значение. Например, погружение в слабый раствор серной кислоты лапки Финальной лягушки, подвешенной на крючке, или щипок кожи конечности пинцетом вызывают отдергивание конечности вследствие сгибания ее в коленном суставе, а при более сильном раздражении — и в тазобедренном суставе. Рефлексы, возникающие Проприорецепторов, могут быть моносинаптическими и поли-Инаптическими, например шейные познотонические рефлексы, азные рефлексы с проприорецепторов участвуют в формировали акта ходьбы. По степени выраженности фазных сгибательных или двигательных рефлексов определяют состояние возбудимости ЦНС и возможные ее нарушения. на сгибательные, а также разгибательные рефлексы возникают при длительном растяжении мышц, их главное назначения поддержание позы. Тоническое сокращение скелетных мышц является фоновым для осуществления всех двигательных актов, осуществляемых с помощью фазических сокращений мышц. В клинике исследуют несколько сгибательных фазных рефлексов: локтевой и ахиллов (проприоцептивные рефлексы), подошвенный (кожный рефлекс). Локтевой рефлекс выражается в сгибании рук в локтевом суставе, возникает при ударе молоточком по сухожилию (при вызове рефлекса рука должна быть слегка согнута в локтевом суставе), его дуга замыкается в пятом-шестом шейных сегментах спинного мозга. Ахиллов рефлекс выражается в подошвенном сгибании стопы в результате сокращения трехглавой мышцы голени, возникает при ударе молоточком по ахиллову сухожилию, рефлекторная дуга замыкается на уровне крестцовых сегментов. Подошвенный рефлекс — сгибание стопы и пальцев при штриховом раздражении подошвы, дуга рефлекса замыкается также на уровне.
Разгибательные рефлексы, как и сгибательные, бывают фазными и тоническими, возникают с проприорецепторов мышц-разгибателей, являются моносинаптическими. Одновременно со сгибательным рефлексом возникает перекрестный разгибательный рефлекс другой конечности.
Фазные рефлексы возникают в ответ на однократное раздражение мышечных рецепторов, например при ударе по сухожилию четырехглавой мышцы ниже надколенной чашечки. При этом возникает коленный разгибательный рефлекс вследствие сокращения четырехглавой мышцы (мотонейроны мышц-сгибателей во время разгибательного рефлекса тормозятся — постсинаптическое реципрокное торможение с помощью вставочных тормозных клеток Реншоу). Рефлекторная дуга коленного рефлекса замыкается во втором—четвертом поясничных сегментах. Фазные разгибательные рефлексы участвуют, как и сгибательные, в формировании акта ходьбы.
Тонические разгибательные рефлексы представляют собой длительное сокращение мышц-разгибателей при длительном растяжении их сухожилий. Их роль — поддержание позы. В положении стоя тоническое сокращение мышц-разгибателей предотвращает сгибание нижних конечностей и обеспечивает сохранение вертикальной естественной позы. Тоническое сокращение мышц спины удерживает туловище в вертикальном положении, обеспечивая осанку человека. Тонические рефлексы на растяжение мышц (сгибателей и разгибателей) называют также миотатическими.
37.
Сухожильный рефлекс возникает на растяжение сухожилия. При сокращении мышцы происходит растяжение сухожилия, в ответ на растяжение сухожилия происходит расслабление мышцы. Рефлекторная дуга состоит из трёх нейронов: мышца – афферентный нейрон спинномозгового узла – афферентный нейрон задних рогов серого вещества СМ – α-мотонейрон передних рогов серого вещества СМ – мышца. Дендрит афферентного нейрона спинномозгового узла образует в сухожилии разветвления оканчивающиеся рецепторами (тельцами Гольджи), аксон – в составе заднего корешка заходит в задние рога серого вещества СМ и образует синаптическую связь с дендритом афферентного нейрона задних рогов серого вещества СМ. Аксон афферентного нейрона задних рогов серого вещества СМ образует синаптическую связь с дендритом α-мотонейрона передних рогов, ответственного за сокращение мышцы, с которой связано сухожилие. Аксон α-мотонейрона выходит из спинного мозга в составе переднего корешка, в составе спинномозгового нерва покидает позвоночный канал, направляется к управляемой мышце, в которой образует многочисленные разветвления. При сокращении мышцы происходит растяжение сухожилия – раздражение телец Гольджи – нервный импульс поступает в афферентный нейрон задних рогов СМ – нервный импульс от афферентного нейрона задних рогов оказывает тормозящее действие на α-мотонейрон – поступление импульсов от α-мотонейрона к мышце прекращается – мышца расслабляется.
38.
Спинальный шок – это изменение мышечного тонуса и отсутствие рефлексов, наблюдающееся сразу после повреждения спинного мозга и связей его с вышележащими структурами ЦНС. Он развивается вследствие выключения влияния вышележащих отделов ЦНС на спинной мозг. Спинальный шок у лягушек длится несколько минут, у собак – несколько дней, у человека – около 2 месяцев. После исчезновения тормозного влияния вышележащих структур ЦНС резко увеличивается тонус мышц, иннервируемых с сегментов спинного мозга ниже места повреждения. Сгибательные и разгибательные рефлексы нижних конечностей резко повышаются. Это происходит вследствие повышения спонтанной активности γ-мотонейронов, приводящей к повышению спонтанной активности мышечных рецепторов и соответственно к повышению тонуса скелетных мышц.
39.
Проводящие пути спинного мозга, составляющие передние, задние и боковые столбы, делятся на восходящие и нисходящие.
Восходящие пути: по ним передается информация от экстеро-, интеро- и проприоцепторов. Тонкий пучок и клиновидный пучок составляют задние столбы спинного мозга – они составлены отростками чувствительных клеток спинальных ганглиев, проводящих возбуждение от проприоцепторов мышц, сухожилий, тактильных рецепторов кожи, висцерорецепторов, и заканчиваются в продолговатом мозге. Передается информация тонко дифференцированной чувствительности о локализации, контуре периферического раздражения и его изменении во времени. Боковой тракт– по нему проводится болевая и температурная чувствительность, а также информация от мышц и суставов к мозжечку.Передний трактотвечает за проведение тактильной чувствительности. Волокна обоих последних трактов идут до таламуса, а далее, после переключения на следующий нейрон, – в кору больших полушарий.Задний и передний спиномозжечковые тракты – образованы аксонами вставочных нейронов задних рогов спинного мозга, по которым информация о позе тела передается в мозжечок. Эта информация необходима для перераспределения тонуса мышц при выполнении движений и сохранения позы и положения тела.
Нисходящие проводящие пути спинного мозга. Пирамидный тракт образован аксонами пирамидных нейронов двигательной зоны коры больших полушарий. По пути от КБП эти аксоны отдают множество коллатералей в структуры промежуточного, среднего, продолговатого мозга и в ретикулярную формацию. В нижней части продолговатого мозга большая часть волокон пирамидного тракта переходит на противоположную сторону, образуя боковой пирамидный тракт в боковом столбе. Другая часть волокон перекрещивается только на уровне того сегмента, на котором аксоны заканчиваются. Обе части волокон пирамидного тракта заканчиваются на мотонейронах передних рогов спинного мозга. Основная роль – управление сокращениями скелетных мышц.
Руброспинальный тракт образован аксонами клеток красного ядра среднего мозга, проходит в боковых столбах, заканчивается на вставочных нейронах спинного мозга. Эти волокна несут информацию от мозжечка, вестибулярного нерва, полосатого тела базальных ганглиев. Роль – управление тонусом мышц и непроизвольной координацией движений.
Вестибулоспинальный тракт образован отростками клеток вестибулярных ядер продолговатого мозга. По нему передается информация от вестибулярного аппарата и мозжечка к мотонейронам, помогающая регулировать тонус мышц, согласованность движений, равновесие.