

72. Шляхи утворення та знешкодження аміаку в організмі.
Головним у кількісному відношенні джерелом наконичення аміаку в організмі людини є окислювальне дезамінування амінокислот, тобто білковий катаболізм: азот сечовини - кінцевого азотовмісного продукту дсградацї білків – складає близько 90 % всього азоту, що екскретується. Додатковими джерелами ендогенного аміаку є реакції дезамінування біогенних амінів, азотистих основ, які утворюються при катаболізмі нуклеотидів. Значна кількість вільного аміаку всмоктується в кров із системи
ворітної вени (v. porta) внаслідок його утворення при катаболізмі азотовмісних біоорганічних сполук (головним чином, білків продуктів харчування) кишковими бактеріями.
Основним джерелом утворення аміаку в тканині головного мозку є реакція гідролітичного дезамінування АМФ до інозинмонофосфату (ІМФ), що каталізується ферментом аденозиндезаміназою:
АМФ+H2O --- ІМФ+NH3
Аміак, що вивільняється, знешкоджується в результаті глуглутамінсинтетазної реакції, утворюючи з L-глутамату глутамін, який виводиться з головного мозку.
Основной реакцией связывания аммиака, протекающей во всех тканях организма, явля ется синтез глутамина под действием глутаминсинтетазы:
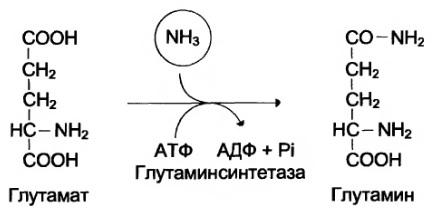
Ещё одной реакцией обезвреживания аммиака в тканях можно считать синтез аспарагина под действием аспарагинсинтетазы.

73. Біосинтез сечовини: послідовність ферментних реакцій біосинтезу, генетичні
аномалії ферментів циклу сечовини.
1. Утворення з аміаку та дюксиду вуглецю за участю АТФ карбамоїлфосфату:
Реакція каталізується карбамоїлфосфатсинтетазою. Джерелом аміногрупи (у вигляді молекули аміаку) с глутаматдегідрогсназна реакція.

2. Перенесення карбамоїльної групи на орнітин з утворенням цитруліну (фермент -
орнітинкарбамоїлтрансфераза):

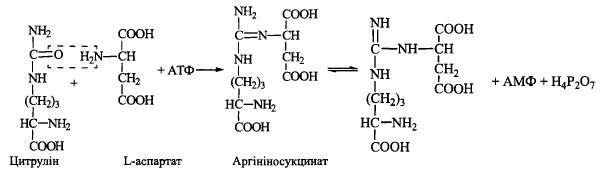
3. Акцептування другої аміногрупи шляхом взаємодії цитруліну з L-аспартатом
(фермент - аргініносукцинатсинтетаза):
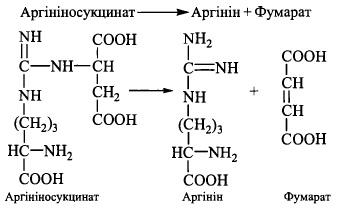
4. Розщеплення аргініносукцинату при дії ферменту аргініносукцинатліази; продуктами реакції є: аргінін - безпосередній попередник сечовини - та фумарат:
5. Гідроліз аргініну при дії ферменту аргінази з утворенням сечовини та регенера-
цією орнітину (завершення метаболічного циклу):

Генетичні дефекти ферментів синтезу сечовини
Існують спадкові ензимопатії, спричинені повним або частковим дефектом утворення в печінці окремих ферментів циклу сечовиноутворення. Найважчими клінічними проявами характеризуються порушення синтезу карбамоїлфосфатсинтетази
та орнітинкарбамоїлтрансферази. Діти з такими генетичними дефектами страждають вираженою енцефалопатією, прояви якої дещо послаблюються в умовах повного виключення споживання харчових білків.
74. Загальні шляхи метаболізму вуглецевих скелетів амінокислот в організмі людини. Глюкогенні та кетогенні амінокислоти.
Двадцять L-амінокислот, що розрізняються за своєю хімічною структурою, біоло-
гічною роллю та особливостями метаболізму, входять до складу білків організму і
присутні в клітинах та екстрацелюлярних просторах у вільному стані. Безазотисті
скелети вільних амінокислот, які утворюються в результаті трансаміпувашія та де-
замінування, - це метаболіти гліколізу, цитратного циклу, Р-окислення жирних кислот
або речовини, що можуть перетворюватися в інтермедіати цих головних катаболіч-
них шляхів організму.
Глюкогенні амінокислоти
L-Амінокислоти, що метаболізуються в циклі трикарбонових кислот, можуть включати свої вуглецеві скелети в молекули глюкози. Ці амінокислоти, використання яких у синтезі глюкози реалізується після їх входження в ЦТК через ацетил-КоА,
а-кетоглутарат, сукциніл-КоА та фумарат, отримали назву глюкогенних амінокислот.
Кетогенні амінокислоти
Дві L-амінокислоти включаються в катаболізм тільки через ацетоацстил-КоА, який
у клітинах печінки може перетворюватися на кетонові тіла ацетоацетат та
β-гідроксибутират. Це - кетогенні амінокислоти. Деякі амінокислоти віддають свої
вуглецеві фрагменти на утворення як глюкози, так і кетонових тіл.
Кетогенез із амінокислот має особливо негативне значення при деяких порушеннях
ферментних процесів, зокрема при некомпенсованому цукровому діабеті, у зв’язку з
чим таким хворим рекомендується обмежувати надходження кетогенних амінокис-
лот у складі продуктів харчування.