

Быков- гистология( общая)
.pdfрые клетки становятся двуядерными. Способность кардиомиоцитов человека к полному митотическому делению утрачивается к моменту рождения или в первые месяцы жизни. Вместе с тем. в этих клетках начинаются процессы полиплоидизации, протекающие, как предполагают, путем обычного, но незавершенного митоза и продолжающиеся в кардиомиоцитах желудочков до 8- 12 лет. Выстраиваясь в цепочки, сердечные миоциты не сливаются друг с другом (как это происходит при развитии скелетного мышечного волокна), а формируют сложные межклеточные соединения - вставочные диски, связывающие кардиомиоциты в функциональные волокна.
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ СЕРДЕЧНОЙ МЫШЕЧНОЙ ТКАНИ
Сердечная мышечная ткань образована клетками - кардиомиоцитами (сердечными миоцитами), связанными друг с другом в области вставочных дисков и образующими трехмерную сеть ветвящихся и анастомозирующих функциональных волокон (рис. 13-11).
Рис. 13-11. Сердечная мышечная ткань Клетки сердечной мышечной ткани - кардиомиоциты (КМЦ) - связаны друг с другом в области вставочных дисков (ВД) и благодаря наличию анастомозов (А) и разветвлений образуют трехмерную сеть функциональных волокон. Ядро (Я) занимает в КМЦ центральное положение, миофибриллы (МФ) располагаются по периферии, под сарколеммой (СЛ). КРС - кровеносные сосуды в прослойках соединительной ткани.
- 431 -
КАРДИОМИОЦИТЫ
Кардиомиоциты - цилиндрические или ветвящиеся клетки, более крупные в желудочках, где их длина составляет 100-150 мкм, а диаметр - 10 - 20 мкм. В предсердиях они обычно имеют неправильную форму и меньшие размеры (длина - 40-70 мкм, диаметр - 5-6 мкм). Кардиомиоциты содержат одно или два ядра и саркоплазму, покрыта сарколеммой, которая снаружи окружена базальной мембраной.
Ядра кардиомиоцитов - светлые, с преобладанием эухроматина, хорошо заметными ядрышками - занимают в клетке центральное положение. У взрослого человека (как и у всех исследованных до настоящего времени млекопитающих) более половины кардиомиоцитов являются двуядерными. Для кардиомиоцитов типична полиплоидия (более выраженная в желудочках), лишь часть из них являются диплоидными (виды с полностью диплоидными сердечными миоцитами не найдены). Степень полиплоидизации кардиомиоцитов характеризуется существенными индивидуальными различиями и даже у молодых здоровых мужчин варьирует в три раза. Предполагают, что степень полиплоидизации кардиомиоцитов у данного индивидуума является важным фактором, определяющим потенциальную способность его сердечной мышцы адаптироваться к повышенным нагрузкам.
Саркоплазма кардиомиоцитов содержит органеллы и включения, которые образуют следующие аппараты: 1) сократительный, 2) передачи возбуждения (с сарколеммы на сократительный аппарат), 3) опорный, 4) энергетический, 5) синтетический, 6) лизосомальный (аппарат внутриклеточного переваривания).
Сократительный аппарат сильно развит в сократительных (рабочих) кардиомиоцитах (в особенности, в желудочковых), которых он занимает до 5070% объема клетки. Слабое развитие этого аппарата свойственно проводящим и секреторным кардиомиоцитам (см. ниже). Сократительный аппарат кардиомиоцитов сходен с таковым в скелетных мышечных волокнах и также представлен миофибриллами, обладающими поперечной исчерченностью (средняя длина саркомера равна примерно 2 мкм). Вместе с тем, миофибриллы кардиомиоцитов нередко частично сливаются друг с другом (рис. 13-12), образуя единую структуру, а их сократимые белки биохимически отличаются от таковых в скелетной мышечной ткани. В саркоплазме кардиомиоцитов миофибриллы ориентированы продольно и располагаются по ее периферии, под сарколеммой.
- 432 -

Рис. 13-12. Саркотубулярная система кардиомиоцита. Саркотубулярная система представлена саркоплазматической сетью (СПС), не образующей терминальных цистерн, и широкими Т-трубочками (Т-ТР). Обозначения компонентов саркомера - те же, что на рис. 13-3. Обратите внимание на частичное слияние МФ друг с другом в единую систему.
Аппарат передачи возбуждения (саркотубулярная система) в
кардиомиоцитах в целом сходен с таковым в скелетных мышечных волоконах, однако он обладает рядом особенностей.
Саркоплазматическая сеть развита слабее, чем в скелетном мышечном волокне, менее активно накапливает Са2+, не образует терминальных цистерн. Во время расслабления она выделяет ионы Са2+ в саркоплазму с низкой скоростью, что обеспечивает автоматизм кардиомиоцитов.
Поперечные (Т-) трубочки - широкие, содержат компоненты базальной мембраны, вместе с элементами саркоплазматической сети образуют диады (включают одну Т-трубочку и одну цистерну сети), которые располагаются в области Z-линий. Т-трубочки хорошо выражены в миоцитах желудочков и почти не обнаруживаются в предсердных миоцитах. Ионы Са2+ проникают в саркоплазму кардиомиоцитов не только из саркоплазматической сети, но также через Т-трубочки и сарколемму из межклеточного пространства.
- 433 -
Опорный аппарат кардиомиоцитов представлен элементами цитоскелета, обеспечивающими упорядоченное расположение миофиламентов и миофибрилл внутри волокна, а также базальной мембраной и сарколеммой. Его структурная и биохимическая организация сходна с таковой в волокнах скелетной мышечной ткани. В кардиомиоцитах его особенностью служит то, что элементы цитоскелета связаны с особыми межклеточными соединениями - вставочными дисками (см. ниже). Сарколемма содержит интегрины - трансмембранные гликопротеины, которые опосредуют связь элементов цитоскелета кардиомиоцитов с компонентами межклеточного вещества (коллагеном, ламинином и фибронектином). Снаружи сарколемма кардиомиоцитов окружена базальной мембраной, в которую вплетаются ретикулярные и тонкие коллагеновые волокна.
Вставочные диски осуществляют связь кардиомиоцитов друг с другом.
Под световым микроскопом они имеют вид поперечных прямых или зигзагообразных полосок, пересекающих функциональное волокно сердечной мышечной ткани (см. рис. 13-11). Под электронным микроскопом определяется сложная организация вставочного диска, представляющего собой комплекс межклеточных соединений нескольких типов (рис. 13-13). В области поперечных (ориентированных перпендикулярно расположению миофибрилл) участков вставочного диска соседние кардиомиоциты образуют многочисленные интердигитации, связанные контактами типа десмосом и полосок слипания (fasciae adherentes). Актиновые филаменты прикрепляются к поперечным участкам сарколеммы вставочного диска на уровне Z-полоски. В области вставочного диска выявляются гликопротеины кадгерины, которые обеспечивают Са-зависимое адгезивное соединение кардиомиоцитов друг с другом. На сарколемме продольных участков вставочного диска имеются многочисленные щелевые соединения, обеспечивающие ионную связь кардиомиоцитов и передачу импульса сокращения.
Энергетический аппарат кардиомиоцитов представлен митохондриями и включениями, расщепление которых обеспечивает получение энергии. Митохондрии лежат рядами между миофибриллами, у полюсов ядра и под сарколеммой. Они очень многочисленные и крупные (занимают около 35-40% объема саркоплазмы - значительно больше, чем в волокнах скелетной мышечной ткани), с плотно расположенными поперечными кристами (см. рис. 13-12), что в совокупности отражает высокий уровень дыхательной активности сердечной мышечной ткани. Митохондрии кардиомиоцитов обладают также свойством накапливать кальций в высоких концентрациях.
- 434 -
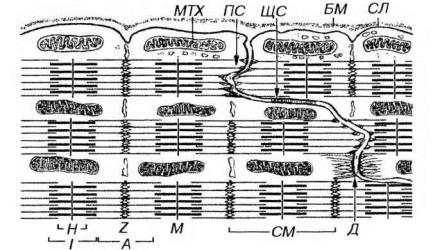
Рис. 13-13. Ультраструктурная организация области вставочного диска кардиомиоцитов. В поперечных участках вставочного диска соседние кардиомиоциты образуют многочисленные интердигитации, связанные контактами типа десмосом (Д). Актиновые филаменты прикрепляются к поперечным участкам сарколеммы вставочного диска в участке полоски слипания (ПС) На сарколемме продольных участков вставочного диска располагаются щелевые соединения (ЩС). БМ - базальная мембрана, СЛ - сарколемма, МТХ - митохондрия. Обозначения компонентов саркомера (СМ) - те же, что на рис. 13-3.
Энергия, необходимая кардиомионитам, получается ими путем расщепления основного энергетического субстрата этих клеток - жирных кислот, переносимых в саркоплазму в форме липопротеинов. Жирные кислоты депонируются в виде триглицеридов в многочисленных липидных каплях, содержание которых значительно выше, чем в скелетных мышцах. Липидные капли располагаются преимущественно у полюсов ядра. Другой энергетический субстрат - гликоген - находится в виде гранул, расположенных преимущественно между миофибриллами и миофиламентами, главным образом, в области I-диска. Кардиомиоциты. как и волокна скелетной мышечной ткани, характеризуются наличием в их саркоплазме железосодержащего кислород-связывающего пигмента миоглобина, придающего им красный цвет и сходного по строению и функции с гемоглобином эритроцитов.
Синтетический аппарат кардиомиоцитов в сократительных (рабочих)
кардиомиоцитах выражен умеренно; он значительно развит лишь в секреторных кардиомиоцитах (см. ниже). Синтетический аппарат включает свободные рибосомы и полисомы, цистерны грЭПС
- 435 -
и элементы комплекса Гольджи, которые располагаются в саркоплазме преимущественно у полюсов ядра.
Лизосомальный аппарат (аппарат внутриклеточного переваривания)
кардиомиоцитов хорошо развит, что отражает высокую скорость обновления их структурных компонентов. Он включает эндосомы, лизосомы и остаточные тельца. Собственно лизосомы располагаются у полюсов ядра и занимают до 10% объема саркоплазмы. Остаточные тельца в виде липофусциновых гранул очень многочисленны (при старении могут составлять до 20% сухой массы миокарда).
Типы кардиомиоцитов
Сердечная мышечная ткань содержит кардиомиоциты трех основных типов: 1) сократительные (рабочие); 2) проводящие 3) секреторные (эндокринные).
1)сократительные (рабочие) кардиомиоциты образуют основную часть миокарда и характеризуются мощно развитым сократительным аппаратом, занимающим большую часть их саркоплазмы (см. выше);
2)проводящие кардиомиоциты обладают способностью к генерации и быстрому проведению электрических импульсов. Они образуют узлы и пучки проводящей системы сердца и разделяются на несколько подтипов. Характеризуются слабым развитием сократительного аппарата, светлой саркоплазмой и крупными ядрами. Особенности распределения и строения различных видов проводящих кардиомиоцитов описаны в курсе частной гистологии (см. раздел "Сердечно-сосудистая система”).
3)секреторные (эндокринные) кардиомиоциты располагаются в предсердиях (в особенности, правом) и характеризуются отростчатой формой н слабым развитием сократительного аппарата. В их саркоплазме вблизи полюсов ядра находятся окруженные мембраной плотные гранулы диаметром 200-300 нм, содержащие гормон - предсердный натриуретический фактор (пептид) - ПНФ (ПНП). Этот гормон вызывает усиленную потерю натрия и воды с мочой (натриурез и диурез), расширение сосудов, снижение артериального давления, угнетение секреции альдостерона, кортизола и вазопрессина. Способностью к выработке ПНФ первоначально в ходе эмбрионального развития обладают все кардиомиоциты; в дальнейшем (уже после рождения) она резко падает в клетках желудочков, сохраняясь в предсердных кардиомиоцитах. При
- 436 -
перегрузке сердечной мышцы способность к синтезу ГТНФ может восстанавливаться в кардиомиоцитах желудочков.
Кровоснабжение сердечной мышечной ткани чрезвычайно обильно: по уровню кровоснабжения (мл/мин/100 г массы) миокард уступает лишь почке и превосходит другие органы, включая головной мозг. В частности, этот показатель для сердечной мышцы в 20 раз выше, чем для скелетной. Сосуды - ветви коронарных артерий - проходят в прослойках соединительной ткани между пучками кардиомиоцитов, распадаясь на капиллярную сеть, в которой каждому миоциту соответствует примерно один капилляр.
Иннервация сердечной мышечной ткани осуществляется волокнами симпатического и парасимпатического отделов вегетативной нервной системы. Она не обусловливает сокращения сердечной мышечной ткани, а лишь регулирует их. Тонкие ветвления нервных волокон с варикозно расширенными участками подходят к кардиомиоцитам, однако не образуют на них нервномышечных окончаний, отделяясь от клеток сравнительно широкой щелью.
РЕГЕНЕРАЦИЯ СЕРДЕЧНОЙ МЫШЕЧНОЙ ТКАНИ
Регенерация сердечной мышечной ткани у взрослого человека может осуществляться только на внутриклеточном уровне путем обновления структурных компонентов кардиомиоцитов, поскольку их способность к пролиферации утрачивается, по-видимому, еще в раннем детстве.
Физиологическая регенерация сердечной мышечной ткани осуществляется на внутриклеточном уровне с высокой интенсивностью, так как для кардиомиоцитов характерна высокая скорость изнашивания и обновления структурных компонентов. Активность этого процесса еще более усиливается при повышенной нагрузке на сердечную мышечную ткань (например, при выполнении тяжелой механической работы, а также в патологических условиях - при гипертонической болезни и пороках сердца). В указанных условиях происходит резко выраженная гипертрофия кардиомиоцитов с увеличением их диаметра до двух раз. При этом нарастают толщина и масса миофибрилл (в которых увеличивается количество саркомеров), а также число митохондрий. В молодом возрасте характерно развитие выраженной полиплоидии кардиомиоцитов.
Репаративная регенерация сердечной мышечной ткани на тканевом и клеточном уровнях у взрослого человека не осуществляется. При
- 437 -
выраженных повреждениях этой ткани (например, в очагах инфаркта миокарда, развивающегося вследствие прекращения кровоснабжения его участка) кардиомиоциты погибают, а на их месте в дальнейшем разрастается соединительная ткань, формирующая рубец. В последние годы установлены новые важные факты о механизмах гибели кардиомиоцитов при инфаркте. Обнаружено, что в очаге инфаркта эти клетки погибают в результате некроза, а в сравнительно широкой зоне, окружающей некротический очаг - механизмом апоптоза. Предполагают, что путем блокирования апоптоза кардиомиоцитов в этой зоне можно уменьшить общие размеры очага повреждения сердечной мышцы.
ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ
Гладкая мышечная ткань очень широко распространена в организме: она входит в состав стенки полых (трубчатых) внутренних органов - бронхов, желудка, кишки, матки, маточных труб, мочеточников, мочевого пузыря (висцеральная гладкая мышечная ткань), а также сосудов (васкулярная гладкая мышечная ткань). Васкулярная гладкая мышечная ткань отличается от висцеральной некоторыми структурными, биохимическими и функциональными особенностями, чувствительностью к действию ряда гормонов, нейромедиаторов и фармакологических препаратов. Гладкая мышечная ткань встречается также в коже, где она образует мышцы, поднимающие волос, а также в капсулах и трабекулах некоторых органов (селезенка, яичко).
Движения, осуществляемые гладкой мышечной тканью, - сравнительно медленные и продолжительные, она обеспечивает также длительные тонические сокращения. Ее сокращения вызывают изменения величины просвета трубчатых органов и лежат в основе их перистальтики. Благодаря сократительной активности этой ткани обеспечивается деятельность органов пищеварительного факта, регуляция дыхания, крово- и лимфотока, выделение мочи, транспорт половых клеток и др.
Помимо собственно гладкой мышечной ткани, развивающейся из мезенхимы (см. ниже), у человека встречаются миоэпителиальные и мионейрапьные клетки, обладающие свойствами гладких миоцитов, но отличающиеся от типичных гладких миоцитов мезенхимного генеза своим происхождением и распределением (ограниченным несколькими четко очерченными участками организма). Первые являются видоизмененными эпителиальными клетками и располагаются в некоторых железах, вторые развиваются из нейрального зачатка и обнаруживаются в радужке глаза. Как уже отмечалось выше, некоторыми авторами эти клетки выделяются в отдельные самостоятельные гистогенетические типы глад-
- 438 -
кой мышечной ткани наряду с наиболее распространенным - мезенхимным.
ГИСТОГЕНЕЗ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ
Развитие гладкой мышечой ткани происходит на сравнительно ранних этапах эмбриогенеза; ее источником служит мезенхима, выселяющаяся из спланхнотомов (образует гладкую мышечная ткань внутренних органов и сосудов) и дерматома (образует гладкую мышечную ткань кожи). По мере дифференцировки клетки удлиняются, в них начинают синтезироваться белки сократительного аппарата и цитоскелета, формируются плотные тельца. В малодифференцированных гладких миоцитах сильно развиты грЭПС и комплекс Гольджи, которые в дальнейшем редуцируются по мере их созревания при одновременном нарастании содержания миофиламетов. Гладкие миоциты продолжают делиться уже после формирования сократительного аппарата, в той или иной степени сохраняя эту способность и в зрелых тканях.
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ
Структурно-функциональной единицей гладкой мышечной ткани мезенхимного типа служит гладкий миоцит (гладкая мышечная клетка).
ГЛАДКИЕ МИОЦИТЫ
Гладкие миоциты - одноядерные клетки преимущественно веретеновидной формы, не обладающие поперечной исчерченностью и образующие многочисленные соединения друг с другом (рис. 13-14 и 13-15). Длина клеток в состоянии расслабления варьирует в пределах 20-1000 мкм (составляя, в среднем, около 200 мкм), их толщина колеблется от 2 до 20 мкм. При резком сокращении длина миоцитов может уменьшаться до 20% начальной. Наиболее крупные клетки характерны для стенки внутренних органов (максимальной длины 500-1000 мкм достигают миоциты матки при беременности), самые мелкие (длиной около 20 мкм) располагаются в стенке сосудов. Гладкие миоциты окружены сарколеммой, которая снаружи покрыта базальной мембраной, содержат одно ядро и саркоплазму, в которой располагаются органеллы и включения.
- 439 -
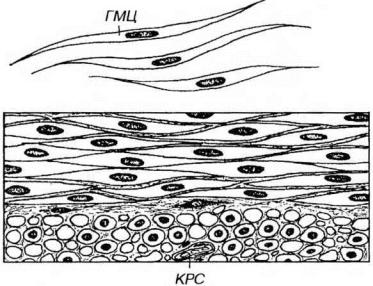
Рис. 13-14. Гладкая мышечная ткань. Сверху показаны изолированные гладкие миоциты (ГМЦ), внизу - их пласт, образованный двумя слоями, в которых клетки ориентированы во взаимно перпендикулярных плоскостях (ГМЦ видны на продольном и поперечном разрезах). КРС - кровеносный сосуд в прослойке рыхлой волокнистой соединительной ткани.
Ядро гладких миоцитов - сигарообразной формы, расположено вдоль длинной оси клетки в ее центральной утолщенной части; при сокращении миоцита оно образует складки и может штопорообразно закручиваться. В расслабленном миоците его длина составляет 10-25 мкм, диаметр - 1-3 мкм. Ядро обычно диплоидное, в нем преобладает эухроматин, выявляются 1-2 ядрышка.
Саркоплазма гладких миоцитов содержит умеренно развитые органеллы общего значения, которые располагаются вместе с включениями в конусовидных участках у полюсов ядра. Периферическая ее часть занята миофиламентами. В саркоплазме выделяют следующие аппараты: 1) сократительный, 2) передачи возбуждения (с сарколеммы на сократительный аппарат), 3) опорный, 4) энергетический, 5) синтетический, 6) лизосомальный (аппарат внутриклеточного переваривания).
Сократительный аппарат гладких миоцитов представлен тонкими
(актиновыми) и толстыми (миозиновыми) филаментами, которые, однако, в отличие от поперечнополосатых мышечных тканей, не формируют миофибрилл.
- 440 -