

Быков- гистология( общая)
.pdfРис. 13-15. Ультраструктурная организация гладких миоцитов. Ядро (Я) располагается в центре клетки и окружено эндоплазмой (ЭПЛ) - участком цитоплазмы, содержащим органеллы общего значения - митохондрии (МТХ), комплекс Гольджи (КГ), цистерны грЭПС. Периферическая часть цитоплазмы (эктоплазма) занята миофиламентами (МФ), связанными с плотными тельцами (ПТ) и плотными пластинками (ПП). Базальная мембрана (БМ) прилежит к сарколемме (СЛ), с которой связаны многочисленные кавеолы (К) В области углублений СЛ по краям миоцита к ней прикреплены коллагеновые фибриллы (КФ). Соседние миоциты образуют интердигитации (ИД) и щелевые соединения
(ЩС).
Тонкие (актиновые) миофиламенты образованы особым набором изоформ актина, свойственным гладким миоцитам, причем помимо мышечного актина в них обнаруживается и немышечный (цитоплазматический) актин. Тонкие филаменты преобладают над толстыми по количеству и занимаемому объему. Они более многочисленны, чем в поперечнополосатых мышечных тканях и располагаются в саркоплазме пучками по 10-20 филаментов, лежащими параллельно или под углом к длинной оси клетки и образующими сетевидные структуры. Концы актиновых филаментов закреплены в особых образованиях, находящихся в саркоплазме или связанных с сарколеммой - плотных тельцах (см. ниже); последние служат также областью фиксации промежуточных филаментов.
- 441 -
Толстые (миозиновые филаменты), в отличие от таковых в поперечнополосатой мышечной ткани, обладают различной длиной (при этом они значительно короче тонких нитей), менее стабильны, не содержат центральной гладкой части, поскольку покрыты миозиновыми головками по всей длине. Это обеспечивает более значительное перекрытие тонких и толстых филаментов, а, следовательно, и большую силу сокращения. Относительное содержание миозиновых филаментов в гладких миоцитах ниже, чем в миофибриллах поперечнополосатой мышечной ткани; на один миозиновый филамент в гладких миоцитах приходится не менее 12 актиновых. По мнению некоторых авторов, миозиновые филаменты гладких миоцитов обладают значительной лабильностью и окончательно собираются непосредственно перед сокращением, распадаясь после него.
Сокращение гладких миоцитов обеспечивается взаимодействием актиновых и миозиновых миофиламентов и развивается в соответствии с моделью скользящих нитей. Оно происходит более медленно и длится дольше, чем в скелетной мышце, что обусловлено более низкой скоростью гидролиза АТФ в гладких миоцитах.
Роль Са2+ в сокращении гладких миоцитов. Как и в поперечнополосатых мышечных тканях, сокращение гладких миоцитов индуцируется притоком Са2+ в саркоплазму, который в этих клетках выделяется саркоплазматической сетью и кавеолами (см. ниже), а также вследствие увеличения проницаемости сарколеммы для данных ионов. Однако (в отличие от поперечнополосатых мышечных тканей) основное влияние Са2+ оказывает не на актиновые, а на миозиновые филаменты. Миозин гладких миоцитов способен взаимодействовать с актином только после фосфорилирования его легкой цепи особым ферментом киназой легких цепей миозина. Этот фермент активируется под влиянием комплекса, образующегося в саркоплазме гладкого миоцита при связывании Са2+ белком кальциймодулином. Дефосфорилирование миозина, происходящее под влиянием фермента фосфатазы миозина, прекращает взаимодействие между актином и миозином и вызывает расслабление гладких миоцитов.
Образование мостиков типа "щеколды" (latch-bridges в англоязычной литературе) является особенностью сократительного аппарата гладких миоцитов: часта миозиновых мостиков после дефосфорилирования не отсоединяется от актина, а остается с ним связанной. Благодаря этому гладкая мышца способна обеспечивать длительное поддержание тонуса без существенных дополнительных энергетических затрат (так как указанные мостики обладают очень медленной циклической активностью).
- 442 -
Регуляция сокращения гладких миоцитов. Активация фермента киназы легких цепей миозина происходит, под действием не только Са2+, но и цАМФ. По этой причине гормоны, повышающие содержание цАМФ в гладких миоцитах, усиливают их сократительную деятельность. Так, эстрогены повышают, а прогестерон снижает уровни цАМФ в гладких миоцитах матки, что обусловливает их сокращение и расслабление, соответственно. Регуляция сокращения гладких миоцитов осуществляется не только на уровне миозиновых нитей, но, отчасти и на уровне актиновых - посредством ряда регуляторных белков: лейотонина (аналога тропонина), кальпонина и кальдесмона.
Опорный аппарат гладкого миоцита представлен его сарколеммой,
базальной мембраной, системой элементов цитоскелета и связанных с ними плотных телец.
Сарколемма каждого миоцита окружена базальной мембраной, в которую вплетаются тонкие ретикулярные, коллагеновые и эластические волокна; коллагеновые фибриллы, прикрепляющиеся к сарколемме в области ее углублений по краям миоцитов, воспринимают усилие, развивающееся при сокращении клеток.
Промежуточные филаменты содержатся в гладких миоцитах в значительно более высоких количествах, чем в кардиомиоцитах и волокнах скелетной мышечной ткани. В гладких миоцитах внутренних органов они образованы преимущественно десмином с небольшим содержанием виментина; в миоцитах сосудов, напротив, основным их компонентом служит виментин, а содержание десмина невелико.
Плотные тельца - овальные или веретеновидные структуры шириной до 0.35 мкм и длиной, в среднем, 1 мкм, лежащие вдоль длинной оси миоцита свободно в его саркоплазме или связанные с внутренней поверхностью сарколеммы (см. рис. 13-15 и 13-16).
Плотные тельца, связанные с сарколеммой, некоторые авторы называют плотными пластинками и считают структурами, не идентичными расположенным в саркоплазме (поскольку они различаются по химическому составу). Более того, в соответствии с современными представлениями, плотные пластинки лишь на срезах кажутся отдельными небольшими образованиями, в реальности же они имеют вид длинных непрерывных "ребер", идущих параллельно друг другу по внутренней поверхности сарколеммы вдоль длинной оси миоцита (см. рис. 13-16).
Плотные пластинки включают периферический и глубокий слои. Первый прилежит к сарколемме и образован филаментами немышечного актина, связанными с трансмембранными белками интегринами по-
- 443 -
средством комплекса адгезивных белков (винкулина, талина, тензина и др.). В глубоком слое филаменты мышечного актина прикреплены к молекулам немышечного актина связующими белками (например, филамином).
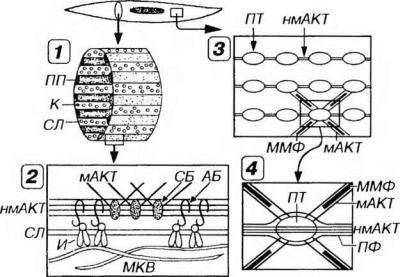
Рис 13-16. Взаимосвязь элементов цитоскелета и сократительного аппарата гладкого миоцита. 1 - плотные пластинки (ПП) в виде "ребер” идут параллельно друг другу по внутренней поверхности сарколеммы (СЛ) вдоль длинной оси миоцита. Кавеолы (К) располагаются в зонах между ПП. 2 - ПП включает периферический и глубокий слои. Первый прилежит к СЛ и образован филаментами немышечного актина (нмАКТ), связанными с трансмембранными белками интегринами (И) посредством комплекса адгезивных белков (АБ). В глубоком слое филаменты мышечного актина (мАКТ) прикреплены к молекулам нмАКТ связующими белками (СБ). МКВ - межклеточное вещество (фибриллы, взаимодействующие с И). 3 - плотные тельца (ПТ) посредством филаментов нмАКТ соединены друг с другом в цепочки, продольно лежащие в саркоплазме; 4 - пучки филаментов мАКТ проникают в ПТ под углом (на фрагменте "З" изображены частично), а промежуточные филаменты (ПФ) окружают их по периферии. Сокращение клетки обеспечивается взаимодействием актиновых и расположенных между ними миозиновых миофпламентов (ММФ) в соответствии с моделью скользящих нитей.
Плотные тельца, свободно лежащие в саркоплазме, согласно новейшим данным, не разбросаны диффузно (как полагали ранее), а располагаются вдоль длинной оси клетки в виде цепочек с интервалом около 2 мкм, соединяясь друг с другом нитями немышечного (цитоплазматического) актина. Пучки филаментов мышечного актина проникают в плотные тельца под углом, а пучки промежуточных филаментов окружают их по периферии (см. рис. 13-16).
- 444 -
Плотные тельца содержат высокие концентрации α-актинина и десмина (в тельцах, связанных с сарколеммой, обнаруживаются также талин и винкулин). Из-за связи с актиновыми филаментами плотные тельца рассматривают как структуры, гомологичные Z-полоскам в поперечнополосатых тканях: их связь с промежуточными филаментами сближает их с пластинками прикрепления десмосом.
Аппарат передачи возбуждения (с сарколеммы на сократительный аппарат) в гладких миоцитах изучен недостаточно. К нему относят саркотазматическую сеть, которая в этих клетках рудиментарна и состоит из системы мелких цистерн и пузырьков, а также особые мембранные структуры - кавеолы. Т-трубочки отсутствуют.
Кавеолы - колбовидные впячивания поверхности сарколеммы диаметром около 70 нм (с более узкой "шейкой"), располженные перпендикулярно длинной оси клетки. Кавеолы открыты в сторону межклеточного пространства, часто располагаются рядами вдоль длинной оси миоцита (занимая промежутки между плотными пластинками), иногда уходят вглубь его саркоплазмы в виде ветвящихся цепочек. Они очень многочисленны (до нескольких сотен тысяч в одной клетке); площадь их суммарной поверхности составляет около 1/3 площади поверхности сарколеммы. Число кавеол не меняется при сокращении, расслаблении или растяжении клетки, они, по-видимому, не участвуют в процессах эцдоцитоза. Кавеолы содержат высокие концентрации кальция, а в их мембране имеются белки, обеспечивающие транспорт кальция в саркоплазму и из нее. Местами они контактируют с элементами саркоплазматической сети. Кавеолы, по-видимому, не только гомологичны системе Т-трубочек поперечнополосатых мышечных тканей, но и выполняют ряд функций, свойственных саркоплазматической сети.
Энергетический аппарат гладких миоцитов представлен митохондриями, а также включениями, содержащими субстраты, расщепление которых обеспечивает энергетические потребности клеток. Митохондрии в гладких миоцитах - сравнительно мелкие, с умеренно развитыми кристами, располагаются вместе с небольшими скоплениями гранул гликогена и мелкими липидными каплями преимущественно у полюсов ядра. Часть митохондрий лежит под сарколеммой. Включения гликогена наиболее многочисленны в миоцитах матки при беременности, содержание липидов особенно велико в васкулярных гладких миоцитах.
Синтетический аппарат гладких миоцитов представлен элементами грЭПС и комплексом Гольджи, лежащими у полюсов ядра,
- 445 -
а также свободными рибосомами, которые располагаются, наряду с этими участками, по всей саркоплазме. Благодаря выраженной синтетической активности гладкие миоциты продуцируют и выделяют (подобно фибробластам) коллагены, эластин и компоненты аморфного вещества. Помимо указанных веществ, они способным синтезировать и секретировать ряд факторов роста и цитокинов. Синтетическая активность гладких миоцитов может резко возрастать в патологических условиях, например, при развитии атеросклеротических изменений в артериях.
Лизосомальный аппарат (аппарат внутриклеточного переваривания)
гладких миоцитов развит сравнительно слабо.
Регуляция сократительной деятельности гладкой мышечной ткани
Сокращение гладкой мышечной ткани происходит под действием нервных импульсов (нейрогенная активность), достигающих сарколеммы миоцитов по эфферентным нервным окончаниям, гуморальных влияний, а также вследствие раздражения миоцитов в отсутствие нервных и гуморальных воздействий (миогенная активность).
Эфферентная иннервация гладкой мышечной ткани осуществляется как симпатическим (норадренергическая иннервация), так и парасимпатическим (холинергическая иннервация) отделами вегетативной нервной системы, которые оказывают противоположное действие на сократительную активность мышечной ткани. Описана также ее серотонинергическая и пептидергическая иннервация. Нервные окончания обнаруживаются лишь на отдельных клетках и имеют вид варикозно расширенных участков тонких веточек аксонов. На соседние миоциты возбуждение передается посредством щелевых соединений.
Афферентная иннервация обеспечивается веточками нервных волокон, образующих свободные окончания в гладкой мышечной ткани (подробнее строение нервных окончаний описано в главе 14).
Гуморальная регуляция активности гладкой мышечной ткани.
Гормоны и другие биологически активные вещества, оказывают влияние на сократительную активность гладкой мышечной ткани (неодинаковое в разных органах) вследствие наличия на ее клетках соответствующих наборов рецепторов. К таким веществам относятся гистамин, серотонин, брадикинин, эндотелии, окись азота, лейкотриены, простагландины, нейротензин, вещество Р, бомбезин, холецистокинин, вазоактиный интестинальный пептид (ВИП), опиоиды и др. Со-
- 446 -
кращения миоцитов матки в конце беременности и во время родов стимулируются гормоном окситоцином - эстрогены повышают, а прогестерон снижает их тонус.
Миогенная активность гладкой мышечной ткани. Физиологическим раздражителем гладких миоцитов служит их растяжение, которое вызывает деполяризацию сарколеммы и приток ионов Са2+ в саркоплазму. Гладкая мышечная ткань характеризуется спонтанной ритмической активностью (автоматией) вследствие циклически меняющейся активности кальциевых насосов в сарколемме. Спонтанная активность наиболее выражена в гладкой мышечной ткани кишки, матки, мочевыводящих путей, она значительно слабее в мышечной ткани кровеносных сосудов. Для автоматии наиболее типичны циклы сокращения и расслабления со средним периодом около 1 мин. (от 0.5 до 2 мин). В обычных условиях на этот миогенный ритм активности влияют нервные и гормональные сигналы, которые усиливают, ослабляют, координируют и синхронизируют сократительную деятельность миоцитов.
РЕГЕНЕРАЦИЯ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ
Физиологическая регенерация гладкой мышечной ткани
осуществляется постоянно на субклеточном уровне путем обновления клеточных компонентов. Регенерации этой ткани на клеточном уровне в физиологических условиях, по-видимому, не происходит (за исключением гладкой мышечной ткани матки при беременности) или, возможно, она осуществляется с очень низкой скоростью. Способность гладкой мышечной ткани к регенерации на клеточном уровне более отчетливо проявляется в условиях повышенных нагрузок.
Гипертрофия гладкой мышечной ткани служит ее реакцией на повышение функциональной нагрузки, обычно связанное с ее растяжением. Гипертрофия этой ткани обусловлена сочетанием процессов гипертрофии и (отчасти) гиперплазии гладких миоцитов. Многократное увеличение массы гладкой мышечной ткани наблюдается как в физиологических условиях (например, в матке при беременности), так и при патологии - в проксимальных участках полых органов при нарушении проходимости их дистальных участков (например, в мочеточнике или желчных путях проксимальнее участка закупорки камнями, в кишке проксимальнее зоны сужения (вызванного опухолью или врожденной аномалией), в мочевом пузыре при нарушении оттока мочи по мочеиспускательному каналу (при аденоме предстательной железы).
- 447 -
Гипертрофия гладких миоцитов проявляется многократным увеличением их размеров, при этом относительное содержание органелл в них, как правило, существенно не меняется. В некоторых органах (например, в сосудах при гипертензии) миоциты могут становиться полиплоидными, однако они всегда остаются одноядерными. Масса коллагеновых волокон, основного вещества и сосудов в мышечной ткани обычно увеличивается пропорционально степени гипертрофии мышечных клеток.
Гиперплазия гладких миоцитов, по-видимому, служит одним из факторов гипертрофии гладкой мышечной ткани, которая развивается при повышенных функциональных нагрузках. Вместе с тем, ее источники остаются предметом дискуссии. Согласно мнению одних исследователей, зрелые гладкие миоциты сохраняют способность к митотическому делению при адекватной стимуляции. Эта возможность отрицается другими авторами, полагающими, что в гладкой мышечной ткани сохраняются малодифференцированные элементы, которые при стимуляции могут превращаться в зрелые гладкие миоциты. Указывают также на возможность преобразования миофибробластов в гладкие миоциты.
Неравномерное разрастание гладкой мышечной ткани наблюдается в патологических условиях и обусловлено различиями чувствительности отдельных гладких миоцитов и их групп к стимулирующим влияниям. Такие изменения нередко наблюдаются в мышечной оболочке матки, в особенности, у женщин старше 30 лет. Они связаны, по-видимому, с нарушением гормональнозависимых процессов регенерации ткани миометрия и проявляются образованием узлов мышечной ткани - миом, имеющих различные размеры и примесь элементов соединительной ткани (в случае выраженности последних такие новообразования носят название фибромиом).
Репаративная регенерация гладкой мышечной ткани развивается после повреждения гладкой мышечной ткани и реализуется за счет тех же источников, что и в нормальных условиях. Способность к полноценному замещению погибшей ткани определяется, по-видимому, объемом повреждения. При достаточно больших зонах повреждения (например, в мышечной оболочке матки после операции кесаревого сечения или в мышечной оболочке кишки после операции создания анастомоза) на месте погибшей гладкой мышечной ткани развивается волокнистая соединительная ткань (вследствие активизации фибробластов соединительнотканных прослоек и дифференцировки расположенных в них малодифференцированных клеток). В этой соединительной ткани постепенно увеличивается содержание коллагеновых волокон, вследствие чего она из рыхлой волокнистой со временем превращается в плотную (рубцовую).
- 448 -
В некоторых случаях усиленная репаративная регенерация в участке повреждения гладкой мышечной ткани нежелательна: после коронарной ангиопластики (операции расширения суженного участка коронарной артерии) спустя некоторое время нередко развивается повторное сужение (рестеноз) вследствие разрастания гладких миоцитов, делящихся под влиянием факторов роста, продуцируемых клетками поврежденной стенки сосуда, в том числе самими миоцитами.
ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ В СОСТАВЕ ОРГАНОВ
В органах гладкая мышечная ткань обычно представлена пластами, пучками и слоями гладких миоцитов. Лишь в отдельных участках (например, в ворсинке тонкой кишки) эти клетки располагаются среди структурных элементов других тканей поодиночке или мелкими группами, не образуя пластов.
Пласты гладких миоцитов образованы очень компактно лежащими клетками, которые разделены промежутками шириной в 40-80 нм (в висцеральной гладкой мышечной ткани) или 200-300 нм (в васкулярной гладкой мышечной ткани). Межклеточные пространства в таких пластах занимают от 10% (в кишке) до 40-50% (в артериях) объема ткани; они заполнены компонентами базальной мембраны, коллагеновыми, ретикулярными и эластическими волокнами, которые в совокупности с отдельными клетками (фибробластами, тучными и малодифференцированными клетками), выявляемыми в более толстых прослойках, образуют эндомизий. Последний содержит сосуды и нервные волокна и способствует объединению гладких миоцитов в пласты и слои. Содержание коллагена в гладкой мышечной ткани в 5-10 раз выше, чем в поперечнополосатых.
Расположение гладких миоцитов в пластах таково, что узкая часть одной клетки прилежит к широкой части другой. Это способствует наиболее компактной укладке миоцитов, обеспечению максимальной площади их взаимных контактов и высокой прочности ткани. В связи с описанным расположением гладких мышечных клеток в пласте на его поперечных срезах соседствуют сечения миоцитов, разрезанных в широкой части и в области узкого края. Формированию пластов гладкими миоцитами способствует образование ими различных связей (по типу миоцит-миоцит, миоцит-клетка другого типа, миоцит-меж-клеточное вещество). В участках межклеточных соединений базальная мембрана миоцитов отсутствует.
- 449 -
Межклеточные соединения гладких миоцитов в пластах обеспечивают механическую и химическую (ионную) связь между ними. К механическим соединениям относят интердигитации и адгезивные соединения, ионную связь осуществляют щелевые соединения.
Соединения типа интердигитации формируются миоцитами, которые часто образуют выросты, вдающиеся в саркоплазму соседних миоцитов.
Адгезивные соединения образованы участками сарколемм двух соседних миоцитов в области расположения связанных с ними плотных телец (плотных пластинок), - мест прикрепления актиновых и промежуточных филаментов - а также специализированным межклеточным веществом между ними. В некоторых участках сарколемма гладких миоцитов формирует аналогичные адгезивные соединения с коллагеновыми и эластическими волокнами (миоцитарно-стромальные соединения).
Плотные соединения между гладкими миоцитами отмечены одними авторами, однако их существование отрицается другими.
Щелевые соединения (нексусы) осуществляют ионную связь между соседними миоцитами, которая обеспечивает распространение возбуждения от клетки к клетке и синхронизацию их сокращений. В области расположения щелевых соединений поверхности соседних миоцитов сближены до расстояния менее 2 нм и связаны друг с другом группами коннексонов различной численности (от 3-6 до 1400), которые занимают до 0.2-0.5% площади поверхности сарколеммы.
Щелевые соединения между гладкими миоцитами при повышении их функциональной активности. Повышение функциональной активности гладких миоцитов - как в физиологических условиях (например, в матке на поздних сроках беременности), так и при патологии (например, в средней оболочке сосудов при повышенном артериальном давлении) - сопровождается резким возрастанием числа и размеров щелевых соединений. В частности, в мышечной оболочке матки число и размеры щелевых соединений, постепенно увеличивающиеся со сроком беременности, особенно резко возрастая непосредственно перед родами, что, вероятно, способствует обеспечению максимальной синхронизации и координации сократительной деятельности миоцитов в процессе родов.
Щелевые соединения между гладкими миоцитами и эндотелиоцитами. В
мелких сосудах гладкие миоциты могут образовывать щелевые соединения не только друг с другом, но и с эндотелиальными клетками. Предполагают, что благодаря связи гладких миоцитов с эндотелиоцитами последние способны влиять на тонус мышечной оболочки сосудов, выделяя факторы, вызывающие сокращение и расслабление миоцитов.
- 450 -