

Быков- гистология( общая)
.pdfНЕРВНЫЕ ВОЛОКНА
Нервные волокна представляют собой отростки нейронов, покрытые глиальными оболочками. Различают два вида нервных волокон - безмиелиновые и миелиновые. Оба вида состоят из центрально лежащего отростка нейрона (осевого цилиндра), окруженного оболочкой из клеток олигодендроглии (в ПНС они называются леммоцитами или шванновскими клетками).
Безмиелиновые нервные волокна у взрослого располагаются преимущественно в составе вегетативной нервной системы и характеризуются сравнительно низкой скоростью проведения нервных импульсов (0.5-2 м/с). Они образуются путем погружения осевого цилиндра (аксона) в цитоплазму леммоцитов, располагающихся в виде тяжей. При этом плазмолемма леммоцита прогибается, окружая аксон, и образует дупликатуру - мезаксон (рис. 14-7). Нередко в цитоплазме одного леммоцита могут находиться до 10-20 осевых цилиндров. Такое волокно напоминает электрический кабель и поэтому называется волокном кабельного типа. Поверхность волокна покрыта базальной мембраной. В ЦНС, в особенности, в ходе ее развития, описаны безмиелиновые волокна, состоящие из "голого" аксона, лишенного оболочки из леммоцитов.
Рис. 14-7. Образование миелинового (1-3) и безмиелинового (4) нервных волокон в периферической нервной системе. Нервное волокно образуется путем погружения аксона (А) нервной клетки в цитоплазму леммоцита (ЛЦ). При образовании миелинового волокна дупликатура плазмолеммы ЛЦ - мезаксон (МА) - наматывается вокруг А, формируя витки миелиновой оболочки (МО). В представленном на рисунке безмиелиновом волокне в цитоплазму ЛЦ погружены несколько А (волокно "кабельного" типа). Я - ядро ЛЦ.
- 471 -
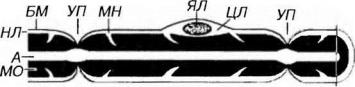
Миелиновые нервные волокна встречаются в ЦНС и ПНС и характеризуются высокой скоростью проведения нервных импульсов (5-120 м/с). Миелиновые волокна обычно толще безмиелиновых и содержат осевые цилиндры большего диаметра. В миелиновом волокне осевой цилиндр непосредственно окружен особой миелиновой оболочкой, вокруг которой располагается тонкий слой, включающий цитоплазму и ядро леммоцита - нейролемма (рис. 14-8 и 14-9). Снаружи волокно также покрыто базальной мембраной. Миелиновая оболочка содержит высокие концентрации липидов и интенсивно окрашивается осмиевой кислотой, имея под световым микроскопом вид однородного слоя, однако под электронным микроскопом обнаруживается, что она возникает в результате слияния многочисленных (до 300) мембранных витков (пластин).
Рис. 14-8. Строение миелинового нервного волокна. Миелиновое волокно состоит из осевого цилиндра, или аксона (А), непосредственно окруженного миелиновой оболочкой (МО) и нейролеммой (НЛ), включающей цитоплазму (ЦЛ) и ядро леммоцита (ЯЛ). Снаружи волокно покрыто базальной мембраной (БМ). Участки МО, в которых сохраняются промежутки между витками миелина, заполненные ЦЛ и поэтому не окрашиваемые осмием, имеют вид миелиновых насечек (МН) МО отсутствует в участках, соответствующих границе соседних леммоцитов - узловых перехватах (УП).
Образование миелиновой оболочки происходит при взаимодействии осевого цилиндра и клеток олигодендроглии с некоторыми различиями в ПНС и ЦНС.
Образование миелиновой оболочки в ПНС: погружение осевого цилиндра в леммоцит сопровождается формированием длинного мезаксона, который начинает вращаться вокруг аксона, образуя первые рыхло расположенные витки миелиновой оболочки (см. рис. 14-7). По мере увеличения числа витков (пластин) в процессе созревания миелина они располагаются все более плотно и частично сливаются; промежутки между ними, заполненные цитоплазмой леммоцита, сохраняются лишь в отдельных участках, не окрашиваемых осмием - миелиновых насечках (Шмидта-Лантермана). При формировании миелиновой оболочки цитоплазма и ядро леммоцита оттесняются к периферии волокна, образуя нейролемму. По длине волокна миелиновая оболочка имеет прерывистый ход.
- 472 -
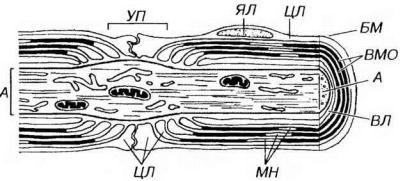
Рис. 14-9. Ультраструктурная организация миелинового нервного волокна. Вокруг аксона (А) располагаются витки миелиновой оболочки (ВМО), снаружи покрытые нейролеммой, а которую входят цитоплазма (ЦЛ) и ядро леммоцита (ЯЛ). Волокно окружено снаружи базальной мембраной (БМ). ЦЛ, помимо нейролеммы, образует внутренний листок (ВЛ), непосредственно прилежащий к А (расположенный между ним и ВМО), она содержится также в зоне, соответствующей границе соседних леммоцитов - узловом перехвате (УП), где миелиновая оболочка отсутствует, и в участках неплотной укладки ВМО - миелиновых насечках (МН).
Узловые перехваты (Ранвье) - участки в области границы соседних леммоцитов, в которых миелиновая оболочка отсутствует, а аксон прикрыт лишь интердигитирующими отростками соседних леммоцитов (см. рис. 14-9). Узловые перехваты повторяются по ходу миелинового волокна с интервалом, равным, в среднем, 1-2 мм. В области узлового перехвата аксон часто расширяется, а в его плазмолемме присутствуют многочисленные натриевые каналы (которые отсутствуют вне перехватов под миелиновой оболочкой).
Распространение деполяризации в миелиновом волокне осуществляется скачками от перехвата к перехвату (сальтаторно). Деполяризация в области одного узлового перехвата сопровождается ее быстрым пассивным распространением по аксону к следующему перехвату, (так как утечка тока в межузловом участке минимальна благодаря высоким изолирующим свойствам миелина). В области следующего перехвата импульс вызывает включение имеющихся ионных каналов и возникает новый участок локальной деполяризации и т.д.
Образование миелиновой оболочки в ЦНС: осевой цилиндр не погружается в цитоплазму олигодендроцита, а охватывается его плоским отростком, который в дальнейшем вращается вокруг него, теряя цитоплазму, причем его витки превращаются в пластинки миелиновой обо-
- 473 -

лочки (рис. 14-10). В отличие от шванновских клеток, один олигодендроцит ЦНС своими отростками может участвовать в миелинизации многих (до 40-50) нервных волокон. Участки аксона в области перехватов Ранвье в ЦНС не прикрыты цитоплазмой олигодендроцитов.
Рис. 14-10. Образование олигодендроцитами миелиновых волокон в ЦНС. 1 - аксон (А) нейрона охватывается плоским отростком (ПО) олигодендроцита (ОДЦ), витки которого превращаются в пластинки миелиновой оболочки (МО). 2 - один ОДЦ своими отростками может участвовать в миелинизации многих А. Участки А в области узловых перехватов (УП) не прикрыты цитоплазмой ОДЦ.
Нарушение образования и повреждение образованного миелина лежат в основе ряда тяжелых заболеваний нервной системы. Миелин в ЦНС может явиться мишенью для аутоиммунного поражения Т-лимфоцитами и макрофагами с его разрушением (демиелинизацией). Этот процесс активно протекает при рассеянном склерозе - тяжелом заболевании неясной (вероятно, вирусной) природы, связанном с расстройством различных функций, развитием параличей, потерей чувствительности. Характер неврологических нарушений определяется топографией и размерами поврежденных участков. При некоторых метаболических расстройствах возникают нарушения образования миелина - лейкодистрофии, проявляющиеся в детстве тяжелыми поражениями нервной системы.
Классификация нервных волокон
Классификация нервных волокон основана на различиях их строения и функции (скорости проведения нервных импульсов). Выделяют три основных типа нервных волокон:
1. Волокна типа А - толстые, миелиновые, с далеко отстоящими узловыми перехватами. Проводят импульсы с высокой скоростью
- 474 -
(15-120 м/с); подразделяются на 4 подтипа (α, β, γ, δ) с уменьшающимися диаметром и скоростью проведения импульса.
2.Волокна типа В - средней толщины, миелиновые, меньшего диаметра,
чем волокна типа А, с более тонкой миелиновой оболочкой и более низкой скоростью проведения нервных импульсов (5-15 м/с).
3.Волокна типа С - тонкие, безмиелиновые, проводят импульсы со сравнительно малой скоростью (0,5-2 м/с).
Регенерация нервных волокон в ПНС
Регенерация нервных волокон в ПНС включает закономерно развертывающуюся сложную последовательность процессов, в ходе которых отросток нейрона активно взаимодействует с глиальными клетками. Собственно регенерация волокон следует за рядом реактивных изменений, обусловленных их повреждением.
Реактивные изменения нервного волокна после его перерезки. В течение 1-й недели после перерезки нервного волокна развивается восходящая дегенерация проксимальной (ближайшей к телу нейрона) части аксона, на конце которой формируется расширение (ретракционная колба). Миелиновая оболочка в области повреждения распадается, тело нейрона набухает, ядро смещается к периферии, хроматофильная субстанция растворяется (рис. 14-11).
В дистальной части волокна после его перерезки отмечается нисходящая дегенерация с полным разрушением аксона, распадом миелина и последующим фагоцитозом детрита макрофагами и глией.
Структурные преобразования при регенерации нервного волокна. Через 4- 6 нед. структура и функция нейрона восстанавливаются, от ретракционной колбы в направлении дистальной части волокна начинают отрастать тонкие веточки (конусы роста). Шванновские клетки в проксимальной части волокна пролиферируют, образуя ленты (Бюнгнера), параллельные ходу волокна. В дистальной части волокна Швановские клетки также сохраняются и митотически делятся, формируя ленты, соединяющиеся с аналогичными образованиями в проксимальной части.
Регенерирующий аксон растет в дистальном направлении со скоростью 3- 4 мм/сут. вдоль лент Бюнгнера, которые играют опорную и направляющую роль; Шванновские клетки образуют новую миелиновую оболочку. Коллатерали и терминали аксона восстанавливаются в течение нескольких месяцев.
- 475 -
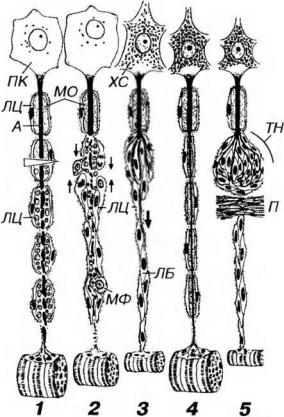
Рис. 14-11. Регенерация миелинового нервного волокна (по R.Krstic, 1985, с изменениями). 1 - после перерезки нервного волокна проксимальная часть аксона (А) подвергается восходящей дегенерации, миелиновая оболочка (МО) в области повреждения распадается, перикарион (ПК) нейрона набухает, ядро смещается к периферии, хроматофильная субстанция (ХС) распадается (2). Дистальная часть, связанная с иннервируемым органом (в приведенном примере - скелетной мышцей) претерпевает нисходящую дегенерацию с полным разрушением А, распадом МО и фагоцитозом детрита макрофагами (МФ) и глией. Леммоциты (ЛЦ) сохраняются и митотически делятся, формируя тяжи - ленты Бюнгнера (ЛБ), соединяющиеся с аналогичными образованиями в проксимальной части волокна (тонкие стрелки). Через 4-6 нед структура и функция нейрона восстанавливаются, от проксимальной части А дистально отрастают тонкие веточки (жирная стрелка), растущие вдоль ЛБ (3). В результате регенерации нервного волокна восстанавливается связь с органоммишенью (мышцей) и регрессирует ее атрофия, вызванная нарушенной иннервацией (4). При возникновении преграды (П) на пути регенерирующего А (например, соединительнотканного рубца) компоненты нервного волокна
- 476 -
формируют травматическую неврому (ТН), которая состоит из разрастающихся веточек А и ЛЦ (5).
Условиями регенерации являются: отсутствие повреждения тела нейрона, небольшое расстояние между частями нервного волокна, отсутствие соединительной ткани, которая может заполнить промежуток между частями волокна. При возникновении преграды на пути регенерирующего аксона формируется травматическая (ампутационная) неврома, которая состоит из разрастающихся аксона и шванновских клеток, впаивающихся в соединительную ткань.
Регенерация нервных волокон в ЦНС отсутствует: хотя нейроны ЦНС обладают способностью к восстановлению своих отростков, этого не происходит, по-видимому, вследствие неблагоприятного влияния микроокружения. После повреждения нейрона микроглия, астроциты и гематогенные макрофаги фагоцитируют детрит в участке разрушенного волокна, на его месте пролиферирующие астроциты образуют плотный глиальный рубец.
НЕРВНЫЕ ОКОНЧАНИЯ
Нервные окончания - концевые аппараты нервных волокон. По функции они разделяются на три группы:
1)межнейронные контакты (синапсы) - обеспечивают функциональную связь между нейронами;
2)эфферентные (эффекторные) окончания - передают сигналы из нервной системы на исполнительные органы (мышцы, железы), имеются на аксонах;
3)рецепторные (чувствительные) окончания воспринимают раздражения из внешней и внутренней среды, имеются на дендритах.
МЕЖНЕЙРОННЫЕ КОНТАКТЫ (СИНАПСЫ)
Межнейронные контакты (синапсы) подразделяются на электрические и химические.
Электрические синапсы в ЦНС млекопитающих редки; они имеют строение щелевых соединений, в которых мембраны синаптически связанных клеток (пре- и постсинаптическая) разделены промежутком шириной 2 нм, пронизанным коннексонами. Последние представляют собой трубочки, образованные белковыми молекулами и служащие водными каналами, через которые мелкие молекулы и ионы могут транспортироваться из одной клетки в
- 477 -

другую (см. главу 3). Когда потенциал действия, распространяющийся по мембране одной клетки, достигает области щелевого соединения, электрический ток пассивно протекает через щель от одной клетки к другой. Импульс способен передаваться в обоих направлениях и практически без задержки.
Химические синапсы - наиболее распространенный тип у млекопитающих. Их действие основано на преобразовании электрического сигнала в химический, который затем вновь преобразуется в электрический. Химический синапс состоит из трех компонентов: пресинаптической части, постсинаптической части и синаптической щели (рис. 14-12). В пресинаптической части содержится (нейро)медиатор, который под влиянием нервного импульса выделяется в синаптическую щель и, связываясь с рецепторами в постсинаптической части, вызывает изменения ионной проницаемости ее мембраны, что приводит к ее деполяризации (в возбуждающих синапсах) или гиперполяризации (в тормозных синапсах). Химические синапсы отличаются от электрических односторонним проведением импульсов, задержкой их передачи (синаптической задержкой длительностью 0.2-0.5 мс), обеспечением как возбуждения, так и торможения постсинаптического нейрона.
Рис. 14-12. Строение химического синапса. Пресинаптическая часть (ПРСЧ) имеет вид концевого бутона (КБ) и включает: синаптические пузырьки (СП), митохондрии (МТХ), нейротрубочки (НТ), нейрофиламенты (НФ), пресинаптическую мембрану (ПРСМ) с пресинаптическим уплотнением (ПРСУ). В постсинаптическую часть (ПОСЧ) входит постсинаптическая мембрана (ПОСМ) с постсинаптическим уплотнением (ПОСУ). В синаптической щели (СЩ) находятся интрасинаптические филаменты (ИСФ).
- 478 -
1.Пресинаптическая часть образуется аксоном по его ходу (проходящий синапс) или представляет собой расширенную конечную часть аксона (концевой бутон). В ней содержатся митохондрии, аЭПС, нейрофиламенты, нейротрубочки и синаптические пузырьки диаметром 20-65 нм, в которых находится нейромедиатор. Форма и характер содержимого пузырьков зависят от находящихся в них нейромедиаторов. Круглые светлые пузырьки обычно содержат ацетилхолин, пузырьки с компактным плотным центром - норадреналин, крупные плотные пузырьки со светлым подмембранным ободком - пептиды. Нейромедиаторы вырабатываются в теле нейрона и механизмом быстрого транспорта переносятся в окончания аксона, где происходит их депонирование. Частично синаптические пузырьки образуются в самом синапсе путем отщепления от цистерн аЭПС. На внутренней стороне плазмолеммы, обращенной к синаптической щели (пресинаптической мембраны) имеется пресинаптическое уплотнение, образованное фибриллярной гексагональной белковой сетью, ячейки которой способствуют равномерному распределению синаптических пузырьков по поверхности мембраны.
2.Постсинаптическая часть представлена постсинаптической мембраной, содержащей особые комплексы интегральных белков - синаптические рецепторы, связывающиеся с нейромедиатором. Мембрана утолщена за счет скопления под ней плотного филаментозного белкового материала (постсинаптическое уплотнение). В зависимости от того, является ли постсинаптической частью межнейронного синапса дендрит, тело нейрона или (реже) его аксон, синапсы подразделяют на аксо-дендритические, аксосоматические и аксо-аксональные, соответственно.
3.Синаптическая щель шириной 20-30 нм иногда содержит поперечно расположенные гликопротеиновые интрасинаптические филаменты толщиной 5 нм, которые являются элементами специализированного гликокаликса, обеспечивающими адгезивные связи пре- и пост-синатической частей, а также направленную диффузию медиатора.
Механизм передачи нервного импульса в химическом синапсе. Под действием нервного импульса происходит активация потенциалзависимых кальциевых каналов пресинаптической мембраны; Са2+ устремляется в аксон, мембраны синаптических пузырьков в присутствии Са2+ сливаются с пресинаптической мембраной, а их содержимое (медиатор) выделяется в синаптическую щель механизмом экзоцитоза. Воздействуя на рецепторы постсинаптической мембраны, медиатор вызывает либо ее деполяризацию, возникновение постсинаптического потенциала действия и образование нервного импульса, либо ее гиперпо-
- 479 -
ляризацию, обусловливая реакцию торможения. Медиаторами, опосредующими возбуждение, например, служат ацетилхолин и глутамат, а торможение опосредуется ГАМК и глицином.
После прекращения взаимодействия медиатора с рецепторами постсинаптической мембраны большая часть его эндоцитозом захватывается пресинаптической частью, меньшая рассеивается в пространстве и захватывается окружающими глиальными клетками. Некоторые медиаторы (например, ацетилхолин) расщепляются ферментами на компоненты, которые далее захватываются пресинаптической частью. Мембраны синаптических пузырьков, встроенные в пресинаптическую мембрану, в дальнейшем включаются в эндоцитозные окаймленные пузырьки и повторно используются для образования новых синаптических пузырьков.
В отсутствие нервного импульса пресинаптическая часть выделяет отдельные небольшие порции медиатора, вызывая в постсинаптической мембране спонтанные миниатюрные потенциалы.
ЭФФЕРЕНТНЫЕ (ЭФФЕКТОРНЫЕ) НЕРВНЫЕ ОКОНЧАНИЯ
Эфферентные (эффекторные) нервные окончания в зависимости от природы иннервируемого органа подразделяются на двигательные и секреторные. Двигательные окончания имеются в поперечнополосатых и гладких мышцах, секреторные - в железах.
Нервно-мышечное окончание (нервно-мышечный синапс, моторная бляшка) - двигательное окончание аксона мотонейрона на волокнах поперечнополосатых соматических мышц - состоит из концевого ветвления аксона, образующего пресинаптическую часть, специализированного участка на мышечном волокне, соответствующего постсинаптической части, и разделяющей их синаптической щели (рис. 14-13).
В крупных мышцах, развивающих значительную силу, один аксон, разветвляясь, иннервирует большое количество (сотни и тысячи) мышечных волокон. Напротив, в мелких мышцах, осуществляющих тонкие движения (например, наружных мышцах глаза), каждое волокно или их небольшая группа иннервируются отдельным аксоном. Один мотонейрон в совокупности с иннервируемыми им мышечными волокнами образует двигательную единицу.
Пресинаптическая часть. Вблизи мышечного волокна аксон утрачивает миелиновую оболочку и дает несколько веточек, которые
- 480 -