

Глава 2 Расово-генетическая компонента цивилизационогенеза
Об актуальности выявления биолого-популяционных оснований цивилизаций и рисках расизма
Феномен биологических популяций раскрывается, как известно, через две основные качественные характеристики. Первая связана с ареалом обитания. Применимость ареальных параметров к сложным социальным системам была доказана в предыдущем разделе. Существование социума неразрывно связано с соответствующей природной средой. Но есть и второй основной параметр характеристики популяций – наследственность. Безусловно, она проявляется на уровне индивидуума. Но значим ли фактор наследственности в применении к социальному существованию? В проверке наличия данной связи состоит целевая установка представленного раздела. В том случае, если гипотеза подтвердится, это будет означать, констатацию биолого-популяционной ипостаси социума. Обнаружение обоих признаков – ареала и наследственности будет указывать на уместность использования инструментария изучения животных популяций к определенным аспектам бытия социальных систем.
Казалось бы, очевидно, что если наследственность проявляется по отношению к каждому человеку – дети похожи на родителей, то она должна проявиться и для их совокупности. Народы объединены общностью происхождения. А это означает их единство не только в социальном плане, но и на генном уровне. Однако тема оказалась политически табуизирована. На этих запретах сказался негативный опыт гипертрофированного преподнесения фактора наследственности.
С одной стороны, это теория биологического происхождения сословного иерархизма. Высшие сословия, согласно ей, антропологически лучшие. Их положение в обществе определяется «благородной наследственностью». Для демократического направления мысли это было категорически неприемлемо.
Другим возможным следствием преувеличенной акцентировки на данном факторе явилась теория расизма. В соответствии с ней наследственность определяет степень социальной успешности рас. Есть высшие и низшие расы. После 1945 г. эта теория была, естественно, выведена за скобки науки.
Но сама-то проблема соотнесения наследственности и специфичности существования сложных социальных систем не была решена. От нее отмахнулись, как от потенциально рискогенной в плане политических выводов. Но не содержит ли таких же рисков абсолютизация и иных факторов развития человечества? Опасность вульгаризации науки, как известно, всегда существовала и очевидно будет существовать. И потому само наличие не есть основание для табуизации тех, или иных сфер познания.
Оба концепта - биологического превосходства социальных групп и рас для нас ценностно неприемлемы. Но это не значит, что фактор наследственности не оказывает влияния на социальные процессы. Оба указанных взгляда являются результатом монистического, уникритериального видения природы социального развития. При универсализации критериев успешности в планетарном смысле возникает, соответственно, иерархия успешных народов. Применение той же логики в рамках одного социума ведет к иерархии социальных кластеров. Однако критерии успешности аксиологически различны. То что значимо для одних народов, может быть менее актуальным для иных. Вариативный подход вместо категорий лучший – худший, оперирует понятием «другой».
Биолого-генетически народы различны. Эти различия находят определенное (причем, не доминантное) преломление в социальной сфере. Базовому для современной биологии понятию генетического кода соответствует понятие цивилизационного кода. Единственно универсальный критерий бытия народов – сама жизнь. Нет такого этноса, который бы исторически (философские концепции могут быть различными) ориентировался на смерть. Но пути повышения жизнеспособности не универсальны. Они задаются в значительной мере биологическо-популяционными различиями человечества. Но далеко не только ими.
Фактор генетического скрещивания
Долгое время в биологии доминировало представление о нескрещиваемости видов. Гибриды, считалось, не способны к производству потомства. Однако еще К. Линней допускал возможность гибридогенного видообразования. Голландский генетик Я. Лотси пошел дальше, рассматривая гибридизацию как основу эволоюционного процесса. Подобно тому, рассуждал он, как индивид имеет двух родителей, так и каждый вид генетически восходит к двум предковым видам. В СССР видным сторонником этого подхода выступал филогенетик Г.М. Попов. Согласно ему, эволюционное развитие следовало бы представлять не в виде традиционного древа, а в форме сети. Каждый из таксонов в ней может взаимодействовать с другими, формируя новый биологический вид. Экспериментально возможность гибридогенного образования была подтверждена в трудах селекционеров – Г.Д. Карпеченко, И.Д. Мичурина, А. Мютцинга, Д.Л. Стеббинса и др. Развитие данного направления теории эволюции привело к констатации существования в природе наряду с описанным Ч. Дарвином дивергенционным, конвергенционного пути создания новых видов. Было установлено то, что видообразование может осуществляться не только в направлении разделения филетических линий, но и их синтеза.
Все эти положения применимы и к вопросу об антропогенезе. Современные исследования генома человека четко фиксируют многочисленные факты межрасовых и межэтнических скрещиваний. На их основе формировались генетически новые человеческие популяции. На биолого-генетической основе выстраивалось далее здание специфичного социального бытия. Возникали этносы. Каждый из них соотносился, как правило, с особым вариантом геномных комбинаций. Культурная традиция находилась в эпоху традиционного общества в интегральном единстве с соответствующим генотипом. Локализованность бытия этносов в этот период определяла генетическую эндогенность, а соответственно, и устойчивость генофонда. Брачная замкнутость нарушалась только в периоды войн и переселений народов. Такого рода миграции вызывали соответствующие волны этногенеза. Процесс глобализации породил новую реальность в геномных изменениях человека. Расширение международных коммуникаций, с одной стороны, объективно разрушает этнические геномы. Брачная изолятивность этносов, хотя и остается значительной в отдельных регионах, в целом по миру уходит в прошлое. С другой стороны, мировые миграции носят теперь преимущественно индивидуальный характер и не выражаются более в формате целенаправленного переселения народов. Отсюда прежняя модель этногенеза, когда скрещивание двух этносов вело к формированию нового генома, более не действует. Генный обмен приобретает открытый в планетарном масштабе характер. В соответствии с этими тенденциями футурологическая перспектива видится в генетической конвергенции человечества. Геномные различия этносов будут со временем все более ослабевать. Конвергенция человечества на биологическом уровне предполагает соотнесение с ней процесса конвергенции цивилизаций.
Применяя к социальным системам модель биологического формообразования, можно предположить, что новые этносы способны возникать как в процессах популяционной дивергенции, так и конвергенции. Дивергенционный процесс связан с адаптацией отдельных групп единого прежде этноса к специфическим средовым условиям бытия. Об этих адаптационных механизмах говорилось в предшествующем разделе. Для нас в данном случае более интересен феномен этнических синтезов. На первый план здесь уже выходит вторая основополагающая характеристика популяций – специфичность их генетической природы. Эта специфика применительно к возникновению этносов достигается посредством вариаций межэтнического скрещивания.
Факт такого синтеза мы обнаруживаем на ранней фазе истории большинства народов. Римская история открывается преданием о похищении первыми римлянами сабинянок, ставшими их женами. Этногенез европейских народов выстраивался в реализованном в раннее средневековье синтезе германского, романского и кельтского компонентов. Русская летописная традиция берет в качестве отправной точки призвание новгородскими славянами варягов. Приведшие к формированию евразийской империи завоевательные походы Чингисхана начались с этнического монгольско-тюрского синтеза, отразившегося в появлении этнонима монголо-татары. Фактор этнического смешения раскрывается при соответствующем анализе как универсальный опыт этногенеза. Это прямо опровергает расистские мифы о «расовой чистоте». Этносы, как новые геномы, исторически возникали в результате популяционных смешений. Генетически чистых, несмешиваемых общностей никогда не существовало. Геномная природа любого народа, по меньшей мере, дуальна. Конечно, в ней может быть доминирующий генный компонент. Но само образование соответствующей этнической группы определялось не этой доминантой, а ее смешением. (См. табл. 2.1).
Табл. 2.1.
Фактор этнического смешения в этногенезе
Название народа |
Участники этногенеза |
|
Племена абазгов, апсилов. |
|
Камиларои, курнаи, нарриньери, вакка, каби, арабана,аранда и др. |
|
Древне германские племена баваров и алеманнов, романизированное население (кельты, реты). |
|
Ирано и тюркоязычные племена, арабы, огузы, печенеги, скифы, булгары, гунны. |
|
Фракийцы, племена иллирийцев, болгары,сербы. |
|
Древние тюркские, самодийские, кетские, угорские племена. |
|
Англичане, индейцы, мексиканцы, африканцы. |
|
Германские племена – англы, саксы, юты, фризы, норманны, бритты. |
|
Мегаэтническая общность, возникшая из соединения аравийских племен с автохтонным населением завоеванных стран. |
|
Испанцы, индейцы, негры, креолы. |
|
Индоевропейцы, хетто-лувийцы, персы, римляне, парфяне, арабы, турки-сельджуки. |
|
Дуррани, гильзаи, вазиры, хоттаки, афридии, шинвари, моманды, хугиани, сафи, какары, оракзаи. |
|
Кыпчаки, аланы. |
|
Финно-угорские племена, ираноязычные сарматы, тюрки, печенеги,булгары, половцы, монголо-татарские племена. |
|
Восточно-славянские племена (дреговичи, кривичи, радимичи, отчасти древлян и волынян); лето-литовские племена (ятвагов). |
|
Бангладешцы, западные бенгальцы, индийцы. |
|
Славянские племена, фракийцы, протоболгары. |
|
Испано-индийские метисы (чоло) и ассимиляции индейцев (каничана, каювава). |
|
Тупи-тапуайя, араваки, караибы, негры, потомки португальцев. |
|
Булагаты, эхириты, хоринцы, хондогоры, эвенки. |
|
Оногуры, протоболгары, палоцы, матьо, куманы, ясы. |
|
Испано-индейские метисы – ладино, креолы. |
|
Саксы, франки, кельты, батавы, фризы. |
|
Протогреческие племена ионийцев, ахейцев, эолийцев, дорийцев, автохтонское население (пеласгов). |
|
Эскимосы, данийцы. |
|
Аджарцы, гурийцы, имеретины, рачинцы, картлийцы, кахетинцы, мохевцы, мтиулы, джавахи,иранцы, ферейданцы, имерхевцы, сваны. |
|
Скандинавские племена (даны), фризы, прибалтийские славяне. |
|
Сефарды, семитоязычные кочевники и земледельцы. |
|
Арабы, местное коптское, берберское и др. населения. |
|
Группа народов, коренное население Америки. |
|
Более сотни этносов, арабы, персы, тюрки. |
|
Яванцы, сунды, мадурцы, малайцы, минангкабау, бугисы, балийцы, банджар, ачех, макассар. |
|
Племена иберов, арабы, берберы. |
|
Италийские племена (италики): латины, этруски, лигуры, венеты, кельты. |
|
Индо-иранские племена, ираноязычные сакские племена, тюркоязычные племена: жуань-жуани, тюрки-тюцзюн, теле, тюргеши, карлуки. |
|
Дербеты, торгуты и другие этнические подразделения ойратов. |
|
Аланы, древние булгары, кыпчаки. |
|
Халаджи, амале, булюки, даррешури, кашкули, фасимадан |
|
Тюркоязычные племена Семиречья и Мавераннахра и карлуков. |
|
Яншаоские племена. |
|
Кастильцы, астурийцы, андалузцы, баски и канарцы, индейцы чибча-муиска. |
|
Три группы племен: палеазиатскиая, алтайская и австронезийская, ведущая роль – протоалтайским племенам. |
|
Иберийские и лигурийские племена, влияние финикийцев, греков, этрусков. |
|
Индейцы аравакского племени сибоней, гуанатахабеи, испанцы, рабы из Африки (йоруба, эве, ашанти и др.). |
|
Тюркоязычные племена( кыпчаки-половцы). |
|
Вьетнамцы, кампучийцы, бирмяне, тайцы. |
|
Балтийские племена, курши, латгалы, земгалы, селы. |
|
Арабские племена, коренное берберское население. |
|
Балтийские племена: аукштайты, жемайты, скальвы, надрувы, а также часть судавов, куршей, земгалов, селов и северных пруссов. |
|
Группы индейских народов: лакандоны, чоли, чонтали, чорти, чухи, цельтали, цоцили, тотики, хакальтеки, хуастеки, киче, какчикели, цутушили, успантеки, маме, ишили, агуакатеки, кекчи, покоман, покомчи. |
|
Славянские племена, местное иллирофракийское население, греки, влахи, сербы. |
|
Выходцы из Индонезии, арабы и арабизированные банту (суахили). |
|
Аборигены: семанги, сенои, джакуны. Выходцы из Индонезии: яванцы, минангкабау, бугисы. |
|
Ацтеки, майя, отоми, миштеки, сапотеки |
|
Волохи и восточные славяне. |
|
Дун-ху, сяньби, хиишивэй, тюркские и тунгусо-маньчжурские племенами. |
|
Финно-угорские племена, тюркские и славянские племена. |
|
Древнегерманское племенное объединение франков, саксов, баваров, алеманов, романизированное кельтское население и реты. |
|
Гунны, тюрки, угорские и энецкие группы. |
|
Ругии, ауганди, грании, раумы, ранрикии, халейги и трёнды. |
|
Аборигены: создатели кобанской культуры) и ираноязычные народы (скифы, сарматы и особенно аланы). |
|
Индейцы-гуарани, испанцы, негры. |
|
Древнеиранские племена, арабы, тюрки, монголы. |
|
Объединения западно-славянских племен: полян, слензан, вислян, мазовшан, поморян. |
|
Иберийские племена(лузитаны), кельты. |
|
Фракийские племена гетов и даков и их романизация в период Римской Империи. |
|
Восточно-славянские племена (поляне, радимичи, вятичи, кривичи, славяне ильменские), финно-угорские и балтийские(жмудь, ливы, латчалы). |
|
Славянские племена (дукляне, тервуняне, конавляне, захлумяне, наречане). |
|
Индоарийцы, ведды, тамилы. |
|
Славяне, даки, фракийцы, тюрки, балты, финно-угоры и т.д. |
|
Славяне, кельты, авары, германцы. |
|
Славянские племена, авары. |
|
Ираноязычные племена: бактрийцы, согдийцы, парфяне, маргианцы, хорезмийцы, парканцы, сако-массагетские племена. |
|
Тюркоязычные племена (кыпчаки, ноган), волско-камские болгары (булгары), финноугоры. |
|
Коренное население Тибета, живут в основном в Китае. |
|
Тюрки-тугю, енесейские кыргызы, уйгуры. |
|
Огузские и туркменские племена, греки, армяне, лазы, курды, узы, печенеги. |
|
Дахи, массагеты,эфталиты, сармато-аланы. |
|
Финно-угорские племена, русские. |
|
Согдийцы, бактрийцы, хорезмийцы, ферганцы, сако-массагетские племена. |
|
Восточно-славянские племена (поляне, северяне, уличи, тиверцы, волыняне), иранские племена. |
|
Индейцы-чарруа, испанцы, итальянцы, французы, |
|
Австронезийцы, филиппинские горные народы, детермолайцы. |
|
Суоми, хямэ, карелы, саво. |
|
Франкские, фризские, саксонские племена. |
|
Кельтские племена (галлы), германские племена (вестготы, бургунды, франки). |
|
Енисейские кыргызы, телеуты, арины, коты, маторы, камасинцы. |
|
Угорские племена смешались с аборигенным населением. |
|
Качичи, шубичи, свачичи, магоровичи. |
|
Арабы, персы, тюрки. |
|
Адыгейцы, кабардинцы, убыхи, ингуши, чеченцы, осетины. |
|
Славяне, кельтское и германское населения; племена: лучане, зличане, дечане, пшоване и др. |
|
Вайнахи и коренное население Северного Кавказа. |
|
Племена индейцев: пикунче и деагитов, испанцы. |
|
Тюркоязычные племена булгар и суваров, ассимилировавшие местные финно-угорское население. |
|
Юкагиры, эскимосы. |
|
Свеи, ёты, готы и другие германские племена. |
|
Базельцы, бернцы. |
|
Пикты, скотты, бритты. |
|
Испанцы, инки, каньяри. |
|
Тунгусо-маньчжурские племена. |
|
Прибалтйско-финские, финно-угорские, северо-германские племена. |
|
Тюркоязычные племена, местное население (эвенки, юкагиры.). |
|
Пуэсские и австронезийские племена, айнойдное население. |
Движущей силой генных трансформаций выступали исторически расовые и этнические миграции. Прослеживается прямая связь между ними и начальной фазой цивилизационогенеза. Цивилизация в Европе начала складываться после перемещения туда мигрантов – земледельцев из Передней Азии, зоны «плодородного полумесяца». Они совокупно составляли не более 10 % европейского населения. Но их приток вызвал утверждение новой земледельческой парадигмы хозяйствования. И именно эта волна миграции, как утверждают генетики, определила формирование основных черт европейского генофонда. Генетически было установлено также, что формирование ранней средиземноморской культуры соотносилось с широким распространением по всей эйкумене Средиземноморья финикийского генотипа. Индоарийские миграции начала II тыс. до н.э. вызвали появление ведической – индийской и авестийской – иранской цивилизации. История становления китайской цивилизации представляла череду вторжений различных по этнической принадлежности кочевых племен. Генетически прослеживается и наличие в цивилизационогенезе Китая европеоидного компонента. Расово смешанным европеоидно-монголоидным определяют историки состав населения древнейших китайских культур – Яншао и Шан-Инь.
Геном древнего эллина сформировался в результате смешения трех этнических компонентов – ионийского, ахейского и дорийского. Ахейцы и дорийцы выступили первоначально по отношению к автохтонному населению в качестве завоевателей. Ахейцами были, в частности, афиняне, дорийцами – спартанцы. Неассимилируемость спартанцев с автохтонами – илотами являлось скорее исключением в рамках общегреческого контекста генетического смешения. На Западе индоевропейская миграция начала II тыс. до н.э. привела к формированию хетской, урартской и эллинской цивилизации. Походы Александра Македонского имели для Ближнего Востока принципиальное значение не только как фактор культурной, но и масштабной генетической трансформации.
Волны завоевательных миграций привели к исчезновению генотипа населения древних цивилизаций – древнеегипетского и древнешумерского. Так, Египет был последовательно завоеван гиксосами, ассирийцами, персами, греко-македонцами, римлянами, арабами, турками и англичанами. Понятно, что исходное геномное основание древнеегипетского цивилизационногенеза оказалось принципиально изменено. Генетики полагают, что наиболее близки к древним египтянам современные копты, составляющие ныне не более 10% населения Арабской республики.
С «исхода» народа Моисея из Египта в Палестину традиционно ведется отсчёт формирования еврейского цивилизационного анклава. В период «рассеяния» религиозные группы евреев придерживались жесткой традиции брачной замкнутости. Это позволило им избежать естественной ассимиляции. Однако генетический обмен все же осуществлялся. Как следствие, возникшие субэтнические группы – евреи-ашкенази и евреи - сеффарды имеют разительные фенотипические различия. Другие иудейские этнические локалитеты – горские евреи, бухарские евреи, феллаши, таты, караимы, крымчаки – и вовсе принадлежат к иным расовым типам. Соответственно, несмотря на общность религии, под единым маркером иудаизма сложились существенно варьирующиеся культурные модели. Не случайно, современная эпоха, наступившая после разрушения Иерусалимского храма, характеризуется в иудаике как «эпоха смешения».
Древние римляне утверждали, что их прямыми предками являлись выходцы из разрушенной ахейцами Трои. Идея реванша над греками поддерживалась ими весь период античности и была передана в средневековье западноевропейским католикам. Ранняя фраза римского цивилизационогенеза определялась многообразием этногенетических компонентов Италии, основные из которых были представлены греческими колонистами, италийцами и этрусками. Этномиграционное содержание периода перехода от античности к средневековью в Европе не нуждается в комментариях. За этим временем в исторической литературе прочно закрепилось понятие «великого переселения народов». Инерция глобальных миграционных сдвигов действовала около пяти столетий. Итогом этих процессов являлось принципиальное переформатирование генетической и цивилизационной карты Европы и значительной части Азии. Римляне, как этногенетический тип исчезли.
Идеология римлян утверждает их прямое преемство от древних ромеев. Однако генетически опровергают этот взгляд, указывая на доминирование в румынском этногенезе дакского компонента. Современные итальянцы в гораздо большей степени лангобарды, нежели римляне. Одним из опеклипов «великого переселения народов» явилось формирование славянской этничности.
Исторически становление исламской цивилизации было связано с арабской миграционной экспансией. Сегодня территория расселения арабов простирается от Марокко на западе до Ирака – на востоке. Понятно, что в каждом из географических локалитетов происходило их генетическое смешение с разнородными группами автохтонов. Генетически значимым было участие арабов в этногенезе испанцев, португальцев, итальянцев, сардинцев. Другая миграционная волна, сыгравшая принципиальное значение в развитии цивилизации ислама, явилась западная турецкая экспансия. На развалинах Византии была воздвигнута Османская империя. Не арабы, а турки выступили на определённом этапе в качестве новой цивилизационнообразующей силы. Сегодня под единым маркером исламской цивилизации существуют де-факто три центра силы, чётко соотносимые с тремя компонентами этничности – арабским – саудитским, тюркским – турецким и индоарийским – иранским.
Современное японское общество, казалось бы, этнически гомогенно. Однако созданию первого общеяпонского государства Ямато предшествовал процесс генетического смешения переселившихся с Корейского полуострова пуеских монголоидных племен с коренным айноидным и австронезийским населением. Переселенцы принесли с собой поливное земледелие, определив формирование жизненных укладов будущей японской цивилизации. Процесс ассимиляции продолжался до XIX века. Окончательно японский генотип сложился только на фазе цивилизационного подъема, связанного с революцией Мэйдзи.
Генезис русской цивилизации соотносится с фиксацией распространения славянского генотипа в VIII-IX вв. в направлении с юго-запада на северо-восток. Фиксируется также активное смешение его с финно-угорским и балтским компонентом. Вместе с тем, генетически не подтверждается сегодня распространенное прежде представление о значительной монголоизации русских под влиянием ордынского ига. Суммарная частота восточно-евразийских гаплогрупп в русских популяциях не превышает 2 %. Выдвинутый в свое время на Западе миф о монголоизации решал вполне определенные политические задачи. Необходимо было дезавуировать в глазах европейцев российскую цивилизацию, связав ее с маркерами «азиатчины» и «варварства».
К тектоническим цивилизационным сдвигом привела, очевидно, начавшаяся с «эпохи великих географических открытий» колониальная экспансия Запада. Но принципиально в контексте рассматриваемой проблематики важно ее соотнесение с геномными трансформациями. Генные изменения фиксируются еще миграциям периода «крестовых - походов». У современных ливанцев генетики четко фиксируют европейские генотипические примеси. После открытия Америки с конца XV века начался незавершенный до сих пор процесс формирования на американских континентах новых геномов. В Южной Америке, где между колонистами – испанцами - португальцами и автохтонным индейским населениям не было жесткой брачной некомплиментарности, этногенез осуществлялся быстрее.
Метисы сегодня являются преобладающим расовым типом на южноамериканском континенте. В Венесуэле, к примеру, их удельный вес достигает 92 % общей численности населения. Новый генотип формируется буквально на глазах. Наряду с исходными европейским и индейским компонентами, определенную роль здесь сыграли негритянский и индийский генетические элементы. Мулаты, к примеру, по сей день численно преобладают в Гвиане, а индийцы в Гайане и Суринаме. Южноамериканский опыт иллюстрирует в данном случае общую модель этногенеза, как проявления этногенетического смешения. И вот уже все отчетливее проявляется факт существования особой цивилизации Латинской Америки. Новое геномное формирование совпадает, таким образом, с процессом цивилизационогенеза. И характерно, что именно Венесуэла – наиболее метисизированная страна континента претендует сегодня на роль цивилизационного лидера. (См. рис. 2.1).
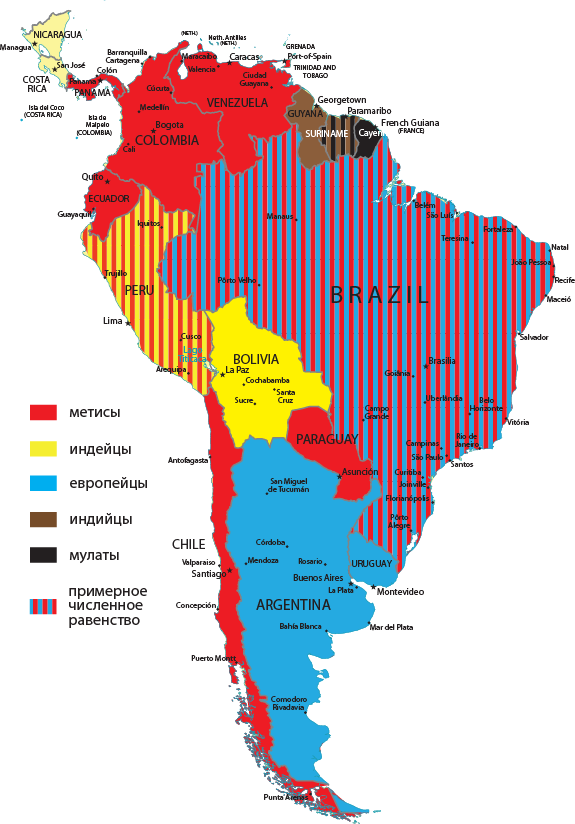
Рис. 2.1. Карта расового доминирования по странам Южной Америки.
В США, как известно, давно был идеологически номинирован образ единого гигантского плавильного этнического котла. Вопреки этой установке в действительности долгое время де-факто там действовали принципы расовой сегрегации. Этнический котел ограничивался главным образом потомками выходцев из Европы. Однако со временем внутрирасовые границы брачности все более размываются. Символом нового этнообразования является фигура действующего президента США Барака Обамы. По линии отца его представляют кенийский народ луо, по матери – англичан, шотландцев, ирландцев, немцев и индейцев чероки. Обамовский геном не поддается этнической идентификации. А между тем, президент воспринимается как персонифицированный образ нации.
Глобализация и многовекторальность миграционных потоков позволяет применительно к сегодняшнему дню говорить о перспективе геномной конвергенции человечества. Уже сейчас в мире количество мигрантов превышает 175 млн. чел., что больше, к примеру, совокупной численности населения России. Но геномная конвергенция произойдет не завтра. Пять из шести жителей планеты проживает еще в том месте, где и родился. Первым в фазу сущностных генетических трансформаций вступит западный мир. Доля мигрантов в нем по отношению к общей численности населения находится в диапазоне от 9,1 % в странах евро и до 20,5 % в Австралии. Для сравнения, в Китае их удельный вес на сегодня составляет лишь пять сотых процента. (См. рис. 2.2.). И миграционное давление на Запад, имея в виду усугубляющиеся цивилизационно-страново диспаритеты в качестве жизни, будет в дальнейшем только усугубляться. Исходя из полученного вывода о зависимости цивилизационогенеза от популяционно-генетических изменений, следует предположить, что грядущая геномная трансформация приведет к существенной трансформации западной цивилизации. Формируется генетически новый тип человека, для которого прежний набор ценностей Запада может оказаться неприемлемым.
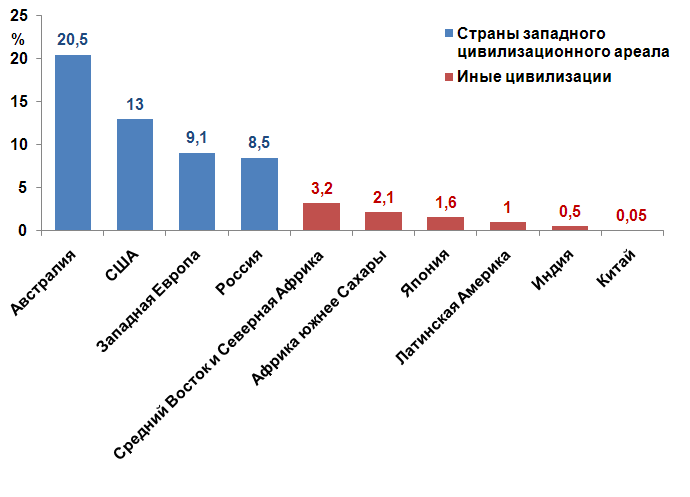
Рис. 2.2. Доля мигрантов в численности населения по ряду цивилизаций и стран мира, в %
О расовом редукционизме
Влияние генно-популяционного уровня жизни на развитие цивилизационных систем, безусловно, существует. Однако действие его факторно ограничено. У А.С. Пушкина, как известно, была по линии деда Ибрагима Ганнибала примесь африканской крови. М.Ю. Лермонтов имел по отцу шотландские корни, а по матери – татарские. Матерью В.А. Жуковского была пленная турчанка. Род Толстых вел свое происхождение от немецкого иммигранта XIV века в Чернигов «мужа честна Индриса», выехавшего «из Цесарския земли». В XVIII веке переехал на службу в Россию из Германии предок А.А. Блока. Отец В.А. Даля был датчанин, мать – француженка. Род Сумароковых ведет свое происхождение по одной версии из Швеции, по другой – из Литвы. Г.А. Державин сам воспел в стихах свое татарское происхождение. Н.М. Карамзин гордился производностью своей фамилии от некого Кара-Мурзы, или Черного Мурзы. К татарской знати принадлежали также предки И.А. Тургенева и И.А. Куприна. Польских предков имели великие русские писатели А.С. Грибоедов, Н.В. Гоголь, Ф.М. Достоевский, Н.А. Некрасов, И.А. Бунин. Данный факт ни коим образом не помешал и Гоголю, и Достоевскому резко высказываться против Польши. Мать А.А. Ахматовой была еврейкой, имевшая к тому же тюркские примеси, восходящие к хану Ахмату. При генной идентификации русская классическая литература вообще переспела бы существовать. Тоже может быть отнесено и к другим сферам бытия. Точно такая же генетическая гетерогенность обнаруживается и в применении к другим национальным культурам.
Вопреки генетике все названные фигуры литераторов составляют лицо русской идентичности. Творчество их внесло сущностный вклад в развитие национальной саморефлексии. Значит дело все- таки не в этнических генах, а в ценностном выборе человека. Это подтверждает выше выдвинутый тезис о приоритетности духовного уровня бытия над биологическим. Неправомочность биолого-популяционного детерминизма следует и из данных самой генетики. Нижеприводимый рисунок иллюстрирует значимость различных уровней принадлежности человека в его геномной структуре. (См. рис. 2.3).
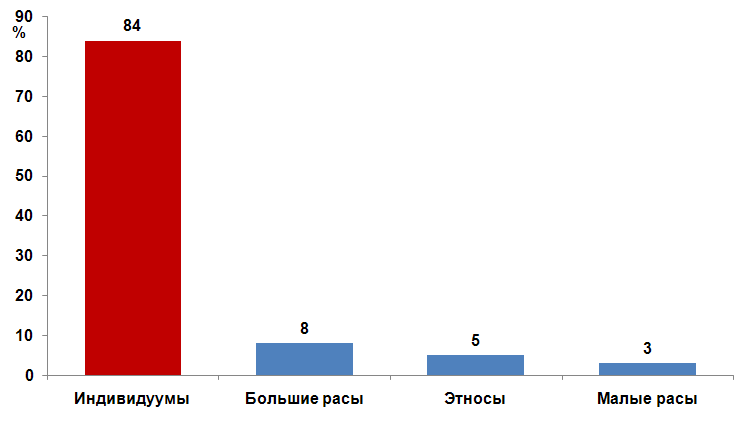
Рис. 2.3. Доля генетического разнообразия в зависимости от расово этнической принадлежности человека, в %
Нельзя сказать, что расовая, или этническая составляющая не играют никакой факторной роли. Но эта факторная роль не превышает совокупно 16%. Индивидумные различия людей оказываются в геноме человека гораздо более значимы. Следовательно, цивилизационогенез расово не детерминирован. Расистский редукционизм, также как и географический, научную неверифицируется.
Используя параметр генетического расстояния, современная генетика позволяет количественно (без расистских мифологем) оценить степень различия рас в общей мегаэволюционной проекции развития мира. Она позволяет также ответить на вопрос - с каким из типов биологических топосов соотносятся сложные социальные системы, такие как цивилизации. Величина расстояния для больших человеческих рас составляет 0,03%. Много это или мало? Для сравнения, межвидовое расстояние составляет от 0,5% выше. Диапозон генетического различия подвидов от 0,17 до 0,22%. Таким образом, не видами, не подвидами расы не являются. Зато показатель генетического расстояния в 0,03% соответствует в животном мире уровню местных популяций. Следовательно, популяция и есть наиболее адекватная категория на уровне биологического измерения, объясняющая процесс расогенеза.
Правильность применения популяционного подхода к пониманию биологического уровня бытия сложных социальных систем подтвердилась. Вместе с тем, была выявлена принципиальная недостаточности сведения объяснительно к парадигме биологии. Это предполагает проследить далее факторную связь биологического и духовного уровней существования человека, предположив влияние в процесс цивилизационогенеза.
Вопросы для обсуждения.
Каково факторное значение расового фактора в цивилизационогенезе в сравнение с иными факторными составляющими?
В чем причины расового разнообразия человечества?
Приведите примеры расово неоднородных цивилизаций. Каковы были выработаны ими механизмы межрасового взаимодействия и расовой толерантности?
Каково влияние географического фактора на формирование расовых различий?
В чем различие между категориями вид и раса?
Каким образом современные открытия в сфере генетики подрывают основы расистского идеологического конструирования?
Каковы социальные корни расизма?
Рекомендуемые темы для докладов и рефератов
Происхождение человечества и география расогенеза.
Исторические вариации расовых классификаций
Расистские законы в национал-социалистской Германии и «лагеря смерти».
Расизм в истории США: феномен ку-клус-кланов.
Геном человека и расогенез.
Феномен апартеида и его историческое крушение.
Международное антирасистское движение.
Раса и национальный темперамент.
Литература
Азимов А. Расы и народы. М, 2005.
Алексеев В.П. География человеческих рас. М., 1974.
Алексеев В.П. Избранное. Т.2. Антропогеография. М.: Наука, 2007.
Алексеев В.П. Историческая антропология. М., 1979.
Алексеева Т.И. Этногенез восточных славян по данным антропологии. М., 1973.
Афонин А.А. Генетика групп крови. М., 2006.
Балановская Е.В., Балановский О.П. Русский генофонд на Русской равнине. М., 2007.
Балановская Е.В., Балановский О.П. Генетические следы исторических и доисторических миграций: континенты, регионы, народы // Вестник ВОГиС. 2009. Т. 13. № 2.
Балибар Э., Валлерстайн И. Раса, нация, класс. Двусмысленные идентичности. М., 2003.
Биология. В 2 кн. / Под ред. В.Н. Ярыгина. М., 2007.
Бунак В.В. Род Homo, его возникновение и последующая эволюция. М., 1980.
Васильев Л.С. Проблемы генезиса китайской цивилизации. М. 1976
Воробьев М.В. Япония в III-VII веках: этнос, общество, культура и окружающий мир. М., 1980.
Голубовский М.Д. Век генетики: эволюция идей и понятий. СПб., 2000.
Гумилев Л.Н. От Руси до России. СПб, 2002.
Гюнтер Г. Избранные работы по расологии. М., 2005.
Дерябин В.Е. Многомерная биометрия для антропологов. М., 1983.
Диас дель Кастильо Б. Правдивая история завоевания Новой Испании. М., 2000
Евдокимов А.Ю. Биосфера и кризис цивилизации. М., 2008.
Животовский Л.А. Популяционная био. метрия. М., 1991.
Киддер Дж. Э. Япония до буддизма: Острова, заселённые богами. М., 2003.
Кинтеро Лопес Р. Встреча цивилизаций или геноцид? // Общественные науки и современность. 1992, № 5;
Назаров В.И. Революция не по Дарвину. Смена эволюционной модели. М., 2011.
Народы мира: историко-этнографический справочник / Гл. ред. Ю.В. Бромлей. М., 1988.
Наследственные болезни в популяциях человека / Под ред. Е.К. Гинтера. М., 2002.
Немировский А. А. У истоков древнееврейского этногенеза. Ветхозаветное предание о патриархах и этнополитическая история Ближнего Востока. М.,2001.
Неструх М.Ф. Человеческие расы. М.: Просвещение, 1965.
Проблема расы в российской физической антропологии / Ред. Т.И. Алексеева, Л.Т. Яблонский. М., 2002.
Рогинский А. А., Левин Н.Г. Антропология: Учебник для студентов ун-тов. М.: Высшая школа, 1978.
Рычков Ю.Г., Ящук Е.В. Генетика и этногенез. Историческая упорядоченность генетической дифференциации популяций человека (модель и реальность) // Вопросы антропологии. Вып. 75. М., 1985.
Сорокина Т.С. Атлас истории медицины: Средние века (476-1640). М., 1983
Стояновский Д.Н. Группа крови и здоровье человека. М., 2004.
Супотницкий М.В., Супотницкая Н.С. Очерки истории чумы. М., 2006.
Чебоксаров Н.Н., Чебоксарова И.А. Народы, расы, культуры. М., 1971.
Юдина Т.Н. Социология миграции. М., 2006.
Coon C.S. The origin of races. New York, 1962.