

Топическая диагностика Нервных болезней
.pdfтысяч нейронов. Большое число синаптических терминалей (синаптических окончаний) лежит с наружной поверх ности тела нейрона, его аксона и дендритов.
Одни из этих синапсов обладают возбуждающим, другие — тормозным воздействием на определенный нейрон (рис. 1.7). Возбуждение всегда распро страняется только в одном направлении: от тела клетки к синапсу. Синапсы по добны переключателям, в которых им пульсы могут быть усилены либо ос лаблены.
Таким образом, нервные клетки по стоянно получают огромное количество сигналов, часть из которых являются возбуждающими, а часть — тормозя щими. В клетке суммируются возбуж дающие импульсы, вычитается сумма тормозящих и затем результат пере дается далее. Импульсы, поступающие по нервному волокну, могут передавать ся через синаптические связи по цепи нейронов в различных направлениях в зависимости от сочетания стимулиру ющих и тормозящих синапсов, встре чающихся на их пути. Поэтому воз действие раздражителя может очень сильно различаться в зависимости от того, какой путь его распространения был выбран.
Центральные влияния
Проприоцепция 9
Проприоцепция
Импульсы, возникающие в мышечных веретенах и сухожильных рецепторах, передаются волокнами типа 1а, наибо лее богатыми миелином и обладающи ми наибольшей скоростью проведения. Другие проприоцептивные импульсы, идущие от рецепторов фасций, суставов и глубоких слоев сединительной ткани, передаются менее миелинизированными волокнами.
Лишь небольшая часть проприоцептивных импульсов достигает коры и, следовательно, осознается. Большинст во же циркулируют в системах обратной связи («обслуживающих» системах), не достигая уровня сознания. Они явля ются элементами рефлексов, которые составляют основу произвольных и про чих движений, а также статических ре флексов, обеспечивающих положение тела в поле действия земного притяже ния.
Периферическая система обратной связи
Прежде чем перейти к обсуждению пу тей болевой, температурной, тактиль ной, вибрационной и прочих видов чув ствительности в спинном и головном
Пирамидный тракт
Рис. 1.9. Круг обратной свя |
|
|
зи при поддержании длины |
|
|
мышцы. |
Клетка Реншоу |
а,-мотонойрон |
10 1 Система чувствительности
мозгу, несколько слов необходимо ска зать о функции различных систем с обратной связью.
На рис. 1.5 показано, что толстое афферентное волокно, идущее от нервномышечного веретена, делится в зоне входа задних корешков в спинной мозг, при этом одна часть волокна вступает в прямой контакт с нейроном переднего рога. Этот нейрон называется мотоней роном, он расположен в сером веществе переднего рога и дает начало эфферент ному волокну. Эфферентные двигатель ные волокна покидают спинной мозг через передние корешки и, миновав спинальный ганглий, на своем пути к скелетной мышце присоединяются к периферическому нерву. Таким обра зом, афферентное и эфферентное во локна образуют дугу, которая идет от мышечного волокна к мотонейрону пе реднего рога и оттуда обратно к скелет ной мускулатуре. Это называется дугой простого моносинаптического рефлекса; она состоит из двух нейронов, соеди ненных посредством синапса.
Моносинаптический проприоцептивный рефлекс
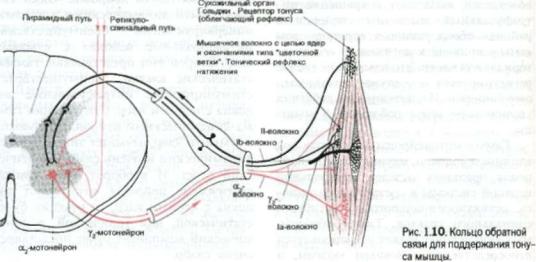
На рис. 1.9 и 1.10 изображены нервномышечные веретена и обслуживаемые ими сухожилия. Каждая мышца содер жит большое количество этих инкапсу лированных, сложных рецепторов рас тяжения, держащих под контролем длину мышцы.
Данные рецепторы состоят из спе циальных мышечных волокон, называ емых интрафузальными волокнами, и из сумко-подобных образований, каж дое из которых в своей центральной, или экваториальной, части содержит 40—50 округлых ядер и называется по этому ядерная сумка. Интрафузальные волокна тоньше, чем обычные экстрафузальные мышечные волокна. Интра фузальные волокна подразделяются на
волокна, ядра которых расположены в сумке, и волокна, ядра которых распо ложены цепью. Эти два типа волокон изображены на рис. 1.9 и 1.10, разде ленными из чисто дидактических целей. В действительности более короткие и тонкие волокна с цепями ядер прикреп ляются к более длинным волокнам, со держащим сумки ядер. В целом вере тено имеет два волокна с ядерной сум кой и четыре — с ядерной цепью.
Как экстрафузальные, так и интра фузальные мышечные волокна иннервируются двигательными волокнами, причем интрафузальными гамма-во локнами (фузимоторными), идущими от небольших мотонейронов, располо женных в передних рогах спинного моз га рядом с большими мотонейронами. Ядерная сумка в каждом веретене оп летена сетью тонких чувствительных нервных волокон, или анулоспиралъных окончаний (по латыни «анулус»означает структуру, имеющую форму спирали). Эти окончания особенно чувствительны к растяжению мышцы. Поэтому вере тена считаются рецепторами натяже ния, ответственными за сохранение мышцей постоянной длины.
Экстрафузальные мышечные волок на в покое имеют определенную длину. Организм всегда старается поддержи вать эту длину постоянной. Как только мышца растягивается, растягивается и веретено. Анулоспиральные нервные окончания немедленно отвечают на рас тяжение мышцы путем генерирования потенциалов действия, которые через быстропроводящие афферентные волок на типа 1а проводятся к большим мо тонейронам спинного мозга, а оттуда, через также быстропроводящие альфа-1 эфферентные волокна, импульс идет к экстрафузалыюй мускулатуре. Так как в результате этого мышца сокращается, то ее первоначальная длина восстанав ливается. Любое растяжение мышцы
незамедлительно запускает в действие этот механизм.
Легкий удар по сухожилию мышцы, например, четырехглавой, на мгновение ее растягивает. Веретена реагируют не медленно. Их импульсы передаются к мотонейрону переднего рога, и он, воз буждаясь, тут же вызывает короткое со кращение мышцы. Этот моносинаптический рефлекс лежит в основе всех проприоцептивных рефлексов. На рис. 1.8 изображены сегменты спинного мозга, участвующие в четырех наиболее важ ных проприоцептивных рефлексах. Очень короткое растяжение мышц, та кое, как вызванное ударами неврологи ческого молоточка, в условиях обычной жизни возникает редко. Механизм об ратной связи для поддержания длины мышцы может быть приспособлен и к обеспечению различной ее длины при воздействии на интрафузальные мы шечные волокна определенных мотор ных систем.
На рис. 1.9. показано, что большие альфа-мотонейроны переднего рога со провождаются более мелкими гамманейронами. Очень тонкие гамма-волок на идут от этих гамма-нейронов к интрафузальным мышечным волокнам. Импульсы, проводимые этими гаммаволокнами, вызывают сокращение интрафузальных мышечных волокон в районах обоих полюсов веретена, тем самым приводя к натяжению его эква ториальной части. Это изменение тотчас регистрируется анулоспиральными окончаниями. Их потенциалы действия увеличивают тонус работающей мыш цы.
Гамма-мотонейроны находятся под влиянием волокон, идущих от мотоней ронов оральных отделов центральной нервной системы в составе пирамидно го, ретикулоспинального и вестибулоспинального трактов. Таким образом, мышечный тонус может регулироваться непосредственно головным мозгом, и
Проприоцепция 11
эта регуляция важна для каждого про извольного движения. Эфферентное воздействие гамма-волокон делает про извольные движения более плавными, «настраивает» их соответственно реша емой задаче. Существует специальная система «гамма-нейрон—мышечное вере тено», которая позволяет через эффе рентное гамма-воздействие регулиро вать реакцию рецепторов растяжения. Сокращение интрафузальных мышеч ных волокон вызывает снижение порога возбудимости рецепторов растяжения; другими словами, даже малое напряже ние мышцы влечет за собой активиза цию рецепторов растяжения. В норме длина мышц автоматически регулиру ется посредством фузимоторной иннер вации через рассмотренную рефлектор ную дугу.
Если первичные и вторичные ре цепторы растягиваются медленно, то в ответ возникает статическая реакция мышечных веретен. Если же растяжение происходит быстро, то ответ является более сильным, динамическим. Как ста тическая, так и динамическая реакции контролируются эфферентными гамманейронами.
Вероятно, существует два типа эф ферентных гамма-нейронов. Один тип представлен динамическими клетками, иннервирующими преимущественно интрафузальные волокна с сумками ядер. Второй тип представляют гаммастатические клетки, преимущественно стимулирующие интрафузальные во локна с цепями ядер. Стимуляция гам ма-динамическими нейронами волокон с сумками ядер вызывает значительный динамический и очень слабый статиче ский ответ. И наоборот, если гаммастатические нейроны возбуждают во локна с цепями ядер, то реакция будет статической, или тонической, а дина мический компонент будет представлен очень слабо.
12 1 Система чувствительности
Другие рефлексы
Быстро проводящие la-нервные волокна проводят потенциалы действия от пер вичных окончаний мышечных волокон с сумками ядер и цепями ядер в цент ральном направлении. Многие мышеч ные веретена, особенно волокна с цепя ми ядер, имеют помимо первичных также вторичные окончания, известные под названием «цветущая ветка». Эти окончания также реагируют на измене ние тонуса, их потенциалы действия проводятся в центральном направлении по тонким II волокнам, связанным с вставочными нейронами, способными осуществлять реципрокное воздействие. Через эти нейроны указанные волокна могут активизировать мышцы-сгибате ли либо мышцы-разгибатели, синхрон но подавляя активность соответствен ных мышц-антагонистов.
На рис. 1.10 показаны мышечные веретена и сухожильный орган (рецеп тор) Гольджи. Эти органы ответственны за напряжение гомонимных мышц, вы званное как активным, так и рефлек торным сокращением; они посылают тормозящие импульсы, которые пере даются через один или два вставочных нейрона. Эти импульсы проводятся по
быстро проводящим волокнам lb типа. Основной задачей рецепторов Гольджи является измерение степени напряже ния конкретной мышцы на основании поступающих в них сигналов и поддер жание мышечного тонуса в физиологи ческих пределах путем посылки тормо зящих импульсов. Таким образом, каж дая мышца находится под контролем двух систем обратной связи: (1) ее дли на контролируется системой, в которой измерительными чувствительными ус тройствами выступают Мышечные ве ретена, и (2) ее тонус контролируется другой системой, в которой измери тельными устройствами являются су хожильные органы Гольджи. Наше тело постоянно подвергается силам гравита ционного воздействия земли. Мы не могли бы стоять вертикально или хо дить, если бы определенные мышцы, такие, как четырехглавая мышца, мыш цы шеи и длинные мышцы спины не противодействовали бы силе гравита ции, имея соответствующий тонус. Од нако в норме уровень этого тонуса не достаточен для поднятия, например, ка кого-то груза. Силы четырехглавых мышц, к примеру, не хватило бы, чтобы не упасть на колени, если бы увеличение
Проприоцепция 13
растяжения мышцы не приводило бы к активизации мышечных веретен, ве дущей, в свою очередь, к мгновенному запуску рефлекса, увеличивающего то нус мышц до необходимого уровня. Благодаря этому механизму происхо дит автоматическое приспособление мышечного тонуса к текущим потреб ностям. Этот же сервисный механизм, основанный на обратной связи, обепе чивает поддержание мышечного тонуса на уровне, необходимом для стояния и ходьбы.
Каждая мышца имеет определенный тонус, так называемый тонус покоя, да же в состоянии полного расслабления. Этот тонус может быть определен при пассивном сгибании или разгибании какойлибо конечности. Для полной ликвидации тонуса необходимо пере сечь все передние корешки, содержащие двигательные волокна к мышцам. Тот же эффект дает пересечение задних ко решков. Таким образом, тонус покоя
создается не самой мышцей, а теми рефлекторными дугами, которые были описаны выше.
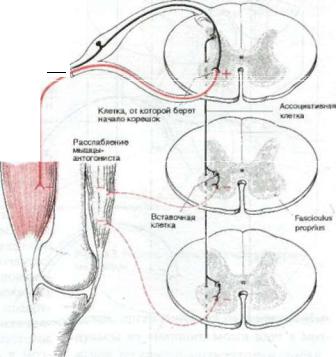
Так называемый моносинаптиче ский рефлекс, строго говоря, не явля ется моносинаптическим. В нем име ется полисинаптический компонент. Для возникновения рефлекторного дви жения конечности необходимо сокра щение главной движущей мышцы, или мышщл-агониста, и синхронное рас слабление противоположной мышцы,
или мышцы-антагониста.
Сокращение, как уже указывалось, является результатом афферентной им пульсации от рецепторов растяжения, возбуждающей мотонейроны передних рогов спинного мозга.
Афферентные волокна, однако, от дают коллатерали к вставочным, или
интернунсиальным нейронам собствен ной нейрональной системы спинного мозга («интернунсио» означает «связу ющий»), и это обеспечивает связь с мо
Анулоспиральный рецептор tffi £
Сокращение мышцы-агониста
Рис. 1.11 Моносинаптический рефлекс и полисинаптическое тормозное воздействие на м ы ш цы-антагонисты.

14 1 Система чувствительности
тонейронами, от вест венными за работу мышц-антагонистов. Вставочные ней роны передают не облегчающие, а тор мозящие импульсы. Этим тормозящим воздействием объясняется тот факт, что
растяжение |
антагонистов, |
|
обусловлен |
|||
ное |
сокращением |
агонистов, |
не вы |
|||
зывает мгновенного |
последующего со |
|||||
кращения |
антагонистов, |
что |
сделало |
|||
бы |
невозможным |
всякое |
движение |
|||
(рис. |
1.11) |
|
|
|
|
|
Другая |
рефлекторная |
дуга |
служит |
|||
инструментом обеспечения |
важного ре |
|||||
флекса сокращения. Это защитный ре флекс, или рефлекс отступления. Он также является полисинаптическим ре
флексом с использованием множества вставочных нейронов, выполняющих роль станций, передающих импульсы. Например, если дотронуться пальцем до горячей плиты, то рука мгновенно отдернется даже до того, как будет по чувствована боль.
В этом примере рецептором явля ется ноцицептор (болевой рецептор). Его потенциалы действия достигают
желатинозной субстанции спинного мозга, где афферентные волокна через синапсы переключаются на многочис ленные вставочные нейроны собствен ной нейрональной системы спинного мозга (клетки путей, вставочные клетки,
ассоциативные клетки, клетки кореш- <ов) — см. рис. 1.12. Начальный имтульс передается ко всем мышцам, уча ствующим в отдергивании руки от объжта, являющегося источником боли. Необходимо обеспечить посылку мно- •очисленных импульсов, реализующих, : одной стороны, сокращение, а с дру- -ой, расслабление соответствующих »1ышц в необходимой последовательногги и с необходимой интенсивностью. Собственная рефлекторная система шинного мозга в чем-то подобна электюнной системе современного компью тера.
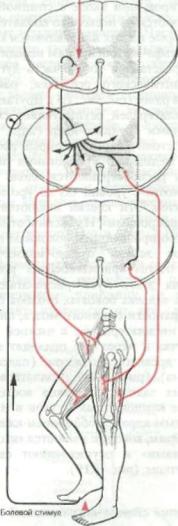
Например, если наступить на ост рый осколок камня, это вызовет боль, соторая немедленно запустит запрог- >аммированную последовательность [вижений (рис. 1.13). Нога, которой фичинена боль, поднимается благодаря тибанию, а на вторую ногу при этом юлностью распределяется вес тела. Внезапное переключение веса привело )ы к падению, если бы мышцы туло1ища, плечевого пояса, шеи и рук не юмпенсировали бы мгновенно наруиенное равновесие и не обеспечили бы •.охранение вертикального положения •ела. Это требует довольно сложных пегемещений сигналов в спинном мозге i их связи с определенными областями :твола мозга и мозжечка. За одной поледовательностью сигналов следует (ругая, определяющая осознание боли, юиск причины, вызвавшей эту боль, смотр ноги с целью выявления ее возложного повреждения.
Большая часть всех этих сигналов >граничивается спинным мозгом. Более 1Ысокие уровни центральной нервной истемы также обычно подключаются ; циркуляции импульсов. В приведенюм примере вовлечение вышележащих ровней было необходимо для предот вращения потери равновесия. Часть имгульсов от мышц, сухожилий, суставов i глубоких тканей направилась в моз-
|
Проприоцепция |
15 |
Кора головного мозга |
/• |
|
Стнол головного мозга |
t |
|
Моэжочок |
|
|
Рис. 1.13. Рефлекс сгибания с полисинаптическими связями.
жечок, орган контроля равновесия. Им пульсы из спинного мозга идут в моз жечок по спинно-мозжечковым путям.
16 1 Система чувствительности
Спиннс—мозжечковые пути
Задний спинно-мозжечковый путь
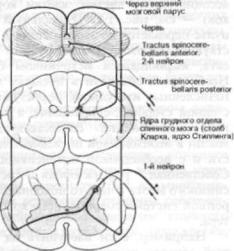
Быстропроводящие la-волокна, идущие от мышечных веретен и сухожильных рецепторов, при входе в спинной мозг разделяются на несколько коллатералей. Некоторые из них направляются к боль шим альфа-мотонейронам передних ро гов, представляя собой часть дуги моносинаптического рефлекса, рассмот ренной ранее (см. рис. 15). Другая груп па коллатералей осуществляет связь с нейронами грудных ядер (ядра Стиллинга, столб Кларка), которые располо жены медиально у основания заднего рога и распространяются вдоль спин ного мозга с уровня С8 до уровня L2сегментов. Эти клетки являются «вто рыми нейронами». Их аксоны относятся
кбыстро проводящим волокнам и фор мируют задний спинно-мозжечковый путь, tractus spinocerebellaris posterior. Волокна этого пути располагаются в задних отделах боковых столбов близко
кповерхности спинного мозга; они вос ходят ипсилатерально к нижней ножке мозжечка и через нее подходят к коре червя древнего мозжечка (палеоцеребеллума) (рис. 1.14). Коллатерали от шейных задних корешков восходят в составе клиновидных пучков к их соб ственным ядрам, добавочным клиновид ным ядрам, которые являются «вторыми нейронами» и осуществляют связь с мозжечком (рис. 1.19).
Передний спинно-мозжечковый путь
Третья группа коллатералей афферент ных la-волокон образуют синапсы с нейронами задних рогов и медиальных отделов серого вещества спинного мозга (рис. 15, 1.14, 1.19). Эти «вторые ней роны», которые расположены по всей длине спинного мозга, включая пояс ничный отдел, дают начало переднему спинно-мозжечковому пути, tractus spi-
Ноосоэнаваемая глубокая чувствительность
Рис. 1.14. Передний и задний спинно-мозжечковые пути.
nocerebellaris anterior. Волокна его вос ходят в передних периферических от делах боковых канатиков как ипсилатеральной, так и контрлатеральной сто роны, и достигают мозжечка. В отличие от заднего спинно-мозжечкового пути, передний путь минует покрышку про долговатого мозга, моста и среднего мозга и входит в червь через верхние ножки мозжечка (соединительные пле чи) и верхний мозговой парус.
Благодаря полисинаптическому проведению импульсов палеоцеребеллум получает информацию о всех аф ферентных сигналах глубокой чувстви тельности и о всех изменениях мышеч ного тонуса. Он контролирует также взаимодействие между мышцами-аго- нистами и антагонистами, необходимое для осуществления стояния, ходьбы и любых других форм движений. Таким образом, на спинальные системы об ратной связи накладываются влияния со стороны более высоких структур, воз действующих на мышцы через экстра пирамидные пути, связи последних с
гамма-мотонейронами передних рогов спинного мозга и эфферентные гаммаимпульсы. Все эти процессы не дости гают уровня сознания.
Задние канатики
Мы осознаем положение наших конеч ностей и состояние тонуса мышц ко нечностей. Мы чувствуем землю под ногами, или, точнее, давление нашего тела на ступни ног. Мы осознаем дви жения в наших суставах. Это свидетель ствует о том, что часть проприоцептивных испульсов достигает коры головно го мозга.
Как уже указывалось выше, проприоцептивная импульсация исходит из рецепторов мышц, сухожилий, фас ций, капсул суставов, глубоких слоев соединительной ткани, кожи. Эти им-
г\ ядрам задних канатиков
Проприоцепция 17
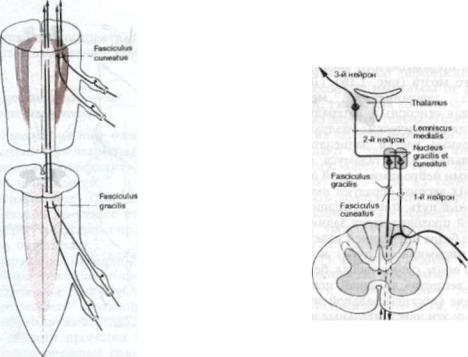
пульсы направляются в спинной мозг по аксонам псевдоуниполярных нейро нов спинальных ганглиев. После того, как аксоны отдают коллатерали к ней ронам передних и задних рогов серого вещества спинного мозга, основная часть центральных ветвей этих аксонов входит в задние канатики. Часть ветвей спускается книзу, а остальные подни маются кверху в составе двух путей, которые в своей совокупности и обра зуют задние канатики спинного мозга: медиально лежащего нежного пучка Голля, fasciculus gracilis, и латерального
клиновидного пучка Бурдаха, fasciculus cuneatus. Эти пути оканчиваются в своих собственных ядрах, нежном ядре, nucleus gracilis, и клиновидном ядре, nucleus cuneatus, расположенных дорсально в покрышке нижних отделов продолговатого мозга (рис. 1.15 и 1.16, см. также рис. 1.19).
Волокна, восходящие в составе зад них канатиков, расположены по соматотопическому принципу. Те, которые проводят импульсы от ног, тазового
Чувство позы, вибрации, давлении, дискриминационная и тактильная чувствительность
Рис. 1.15. Задний канатик. |
Рис. 1.16. Задний канатик. |
18 1 Система чувствительности
пояса и нижней части туловища, идут в нежном пучке, вплотную к задней сре динной перегородке. Волокна, проводя щие импульсы от верхнего плечевого пояса, рук и шеи идут в составе кли новидного пучка, причем ветви от шеи расположены наиболее латерально (см. рис. 1.15). Нервные клетки нежного и клиновидного ядер представляют собой «вторые нейроны». Их аксоны форми руют бульботаламический путь, tractus bulbothalamicus, и связаны с «третьими нейронами», находящимися в заднела-
тералъном вентральном ядре таломуса.
Начинаясь в нежном и клиновидном ядрах, бульботаламические пути идут вначале по передней поверхности про долговатого мозга в его нижних отделах по обе стороны от так называемого центрального серого вещества непосред ственно над перекрестом нисходящих пирамидных путей. Затем они в составе медиальной петли, lemniscus medialis, пе ресекают среднюю линию (перекрест медиальной петли) и восходят к таламусу кзади от пирамид и медиальнее нижних олив, минуя покрышку верхних отделов продолговатого мозга, моста и среднего мозга (рис. 1.16). (Для более легкого запоминания: медиальная петля для се.нсорных, латеральная пет ля — для акустических раздражителей).
Нервные клетки заднелатеральных вентральных ядер таламуса являются «третьими нейронами» в этой цепи (рис. 1.16). Их аксоны образуют таламокортикальный путь, tractus thalamocorticalis, который проходит через задние отделы заднего бедра внутренней капсулы (кза ди от пирамидного пути) и через лу чистый венец, corona radiata, белого моз гового вещества к задней центральной извилине (постцентральной извилине). Поэтому эти чувствительные импульсы являются осознаваемыми. Соматотопическое расположение указанных воло кон на их пути к таламусу и к коре соответствует наблюдаемому в спинном
мозгу. В коре головного мозга чувстви тельная проекция тела соответвует схе ме тела человека, стоящего на голове (рис. 820а). Задние канатики проводят преимущественно импульсы, возни кающие в проприоцепторах и рецепто рах кожи. При их повреждении стано вится невозможным определение поло жения конечностей в пространстве, уз навание предметов на ощупь. Не узна ются цифры и буквы, написанные на коже. Невозможно различение двух од новременно наносимых на кожу раздра жений. Уменьшается восприятие чувст ва давления; поэтому нарушается ощу щение опоры стоп на землю и возникает атаксия при стоянии и ходьбе, в осо бенности в темноте или при закрывании глаз. Эти нарушения характерны для поражения задних канатиков спинного мозга. При поражении нежного и кли новидного ядер, медиальной петли, та-
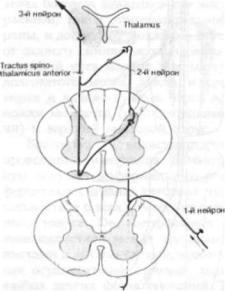
Грубая тактильная чувствительность, мало дифференцированное чувство давления
Рис. 1.17. Передний спинно-таламический путь.