

Топическая диагностика Нервных болезней
.pdf3-й нейрон \
-W Таламус
2-й нейрон
Tractus spinothalamicus lateralis
Substantia gelatinosa 141 нейрон
Боль, температура (чувство щекотки, зуда, сексуальные ощущения)
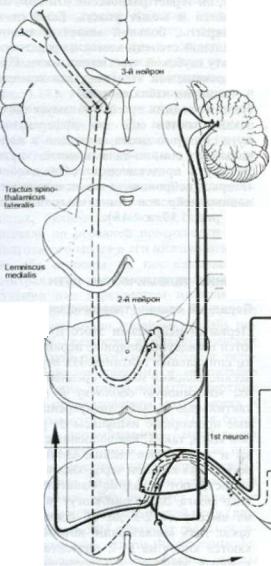
Рис. 1.18. Латеральный спинно-таламический путь.
ламуса или постцентральнои извилины данные проявления менее выражены.
Синдромы поражения задних канатиков
1.Утрата чувства положения и локомоции: При закрытых глазах пациент не может определить положение сво их конечностей.
2.Астереогноз: При закрытых глазах больной не может узнать и описать форму и материал предмета, который он ощупывает.
3.Нарушение двумерной дискриминаци онной чувствительности.
4.Нарушение вибрационной чувстви тельности: Больной не ощущает вибрацию камертона, установленного в области проекции кости.
5.Положительный симптом Ромберга:
Если больной закрывает глаза и ста новится в позу, когда ступни ног ус-
Спинно-тзламические пути |
19 |
танавливаются вплотную друг к дру гу, он теряет равновесие. Он покачи вается и может упасть. Если глаза открыты, больной может в значи тельной степени компенсировать ут рату глубокой чувствительности. При мозжечковой атаксии такая компен сация не наблюдается.
Помимо задних путей, в таламусе также заканчиваются еще два афферентных пути спинного мозга, передний и лате ральный спинно-таламические пути. Эти пути представляют собой аксоны «вторых нейронов», а не псевдоунипо лярных нейронов спинальных ганглиев (см. рис. 1.17 и 1.18).
Спинно-таламические пути
Передний спинно-таламический путь
Первыми нейронами этого пути явля ются псевдоуниполярные нервные клет ки спинальных ганглиев. Их перифери ческие волокна имеют средней толщи ны миелиновую оболочку и проводят тактильные и слабо дифференцирован ные прессорные импульсы от рецепто ров кожи, таких, как волосяные мешоч ки и тактильные тельца. Центральные ветви этих аксонов через задние кореш ки вступают в задние канатики спин ного мозга. Здесь они могут подняться на высоту от 2-х до 15 сегментов, а также дать коллатерали, которые спус каются книзу на 1—2 сегмента. На не которых уровнях эти волокна имеют синапсы с нейронами задних рогов (рис. 1.17). Нейроны задних рогов яв ляются «вторыми нейронами», участву ющими в формировании переднего спинно-таламического пути, tractus spinothalamicus anterior. Этот путь перехо дит на противоположную сторону в сос таве передней спайки, commissura anterior, кпереди от центрального спин номозгового канала и располагается да-
3 — 1873
20 1 Система чувствительности
Tractus spinocerebellaris posted сч
Tractus spi пе
сет ebellaris anterior
Tractus - spino thalamicus anterior
PaleocerebeHum
Tractus sptnoco- |
jГ о м о н м м 1 ш и |
robellaris posterior |
|
Tractus spinoco- |
проприоцопция |
robollaris anterior |
|
Fibrae arcuatao extemae dor sales
Nucleus graalis ot cuneatus
Nucleus cuneatus accessorius
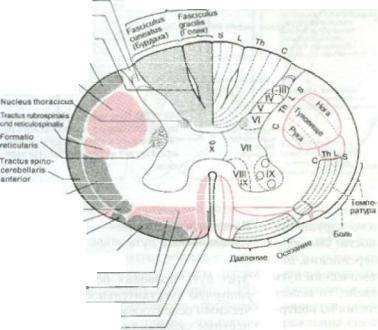
Глубокая чувствительность
мышечные веретена, мышечные и сухожильные роцешоры, рецепторы сусгавови т. д)
Вибрационная, дискриминационная, тактильная чувствительность, чувство положения, давления, осязания
(рецепторы кожи, мышц и сухожилий, тельца ОЭатер-Лачини)
Давление,осязание
(волосяные мешочки и различные рецепторы кожи)
Боль, температура (свободные нораныо окончания, тельца Крауээ и Руффини?)
Рис. 1.19. Наиболее важ ные восходящие пути спинного мозга.
лее в передней периферической зоне переднебоковых канатиков. Отсюда он восходит к заднелатеральному вент ральному ядру таламуса вместе с лате ральным спинно-таламическим путем и медиальной петлей (см. рис. 1.19). Нейроны таламуса являются «третьими нейронами», проводящими импульсы по волокнам таламокортикалыюго пути
к постцентральной извилине коры го ловного мозга. Тот факт, что централь ные ветви первых нейронов поднима ются и спускаются по задним канати кам и связаны многочисленными коллатералями со «вторыми нейронами», является обьяснением тому, что пов реждение спинно-таламического пути в поясничном и грудном отделах обычно

Gyrus
. - postcentralis
Corpus
geniculum laterale
Nucleus ventralis posterolateralis
Lemniscus medialis
Tractus spinothalamicus lateralis
Спинно-таламические пути |
21 |
Плечо Надплечье Голова Шея
- - Туловище Бедро
Верхняя часть ноги
Нихияя часть ноги
Пальцы сюпы, гениталии
Tractus corticospinalis
Lemniscus meoTalis
Tractus spinothalamicus lateralis
Nucleus anterior thalami
"V Таламус (пра вый, переднебоковая проекция)
Рис. 1.20. Путь медиальной петли и ла терального спинно-таламического пу ти к таламусу и далее через внутрен нюю капсулу к постцентральной изви лине.
не вызывает значительного нарушения тактильной чувствительности. Импуль сы легко обходят зону повреждения. Ес ли передний спинно-таламический путь поражается в шейном отделе, то может выявляться легкая гипестезия на контр латеральной ноге.
Латеральный спинноталамический путь
Этот путь проводит болевую и темпе ратурную чувствительность. Перифери ческими рецепторами служат свободные нервные окончания кожи. Они являют-
22 |
1 Система |
чувствительности |
|
|
|
|
|
|
|||
ся окончаниями периферических ветвей |
периферии, а проводящие импульсы от |
||||||||||
псевдоуниполярных нейронов |
спиналь- |
шеи — наиболее медиально (см. рис. |
|||||||||
ных ганглиев; эти ветви относятся к |
121). |
|
|
|
|
||||||
тонким волокнам группы А и почти |
В стволе мозга латеральный спин |
||||||||||
безмиелиновым волокнам группы С. |
но-таламический |
путь присоединяется |
|||||||||
Центральные ветви входят в спинной |
в виде спинальной петли к медиальной |
||||||||||
мозг через латеральные порции задних |
петле. Он оканчивается в вентральных |
||||||||||
корешков. В спинном мозге они разде |
заднелатеральных |
ядрах |
таламуса |
(см. |
|||||||
ляются на короткие продольные колла- |
рис. 5S. VPL и V. с. е.). Отсюда «третьи |
||||||||||
терали, |
которые, |
поднявшись |
на 1 — 2 |
нейроны» |
формируют |
таламокорти- |
|||||
сегмента, образуют синапсы с нейрона |
кальный путь, который направляется к |
||||||||||
ми желатинозной |
субстанции |
(Ролан |
коре задней центральной извилины (см. |
||||||||
да), substantia gclatinosa. Это «вторые |
рис. 1 . 20) . Волокна, проводящие боле |
||||||||||
нейроны», |
образующие |
латеральный |
вую и температурную чувствительность, |
||||||||
спинно-таламический путь, tractus spi |
располагаются в спинно-таламическом |
||||||||||
nothalamicus lateralis (рис. 1 . 18) . Его во |
пути настолько вплотную друг к другу, |
||||||||||
локна переходят через переднюю спайку |
что их невозможно разделить анатоми |
||||||||||
на противоположную сторону и подни |
чески. Поэтому при повреждении лате |
||||||||||
маются в латеральных отделах боковых |
рального |
спиноталамического |
пути |
||||||||
канатиков к таламусу. Как и волокна |
страдает как болевая, так и температур |
||||||||||
задних канатиков, волокна обоих снин- |
ная чувствительность, хотя не всегда в |
||||||||||
но-таламических путей имеют сомато- |
равной степени. |
|
|
|
|||||||
топическую |
организацию: |
идущие |
от |
Латеральный |
спинно-таламический |
||||||
нижних |
конечностей располагаются |
на |
путь является главным |
проводником |
|||||||
Substantia gclatinosa |
|
Funiculus posterior |
|
|
|
|
|
|
|
||
Tractus dorsolaterals |
|
|
|
|
|
|
|
|
|
||
(Lissaucr-Traklus) |
|
|
|
|
|
|
|
|
|
|
|
Tractus semilunaris (Schullzsches Komma)
Trnctus soinocerebellaris postorior
Tractus corticospinalis loloralis
Tractus spino thalamicus lateralis Tractus spinotoctalis Tractus olivospinal Tractus spino-olivaris
Tractus spinothalamicus anterior |
|
|
Tractus vcstibulospinalis |
Рис. 1.21. Срез спинного |
|
Tractus reticulospinal! |
||
мозга с указанием топогра |
||
Tractus tectospinalis |
фии восходящих и нисходя |
|
Tractus corticospinalis anterior |
щих путей и цитоархитекто- |
|
Fasciculus sulcomarginal! |
ники (по Rexed; 1954). |
|
|
болевой и температурной чувствитель ности. При пересечении этого пути во время операции, выполняемой билате рально для устранения боли, неподда ющейся никакому другому лечению (хордотомия), боль может исчезнуть не полностью. Это свидетельствует о том, что болевые импульсы могут прово диться также через вставочные нейроны по собственным волокнам внутренних путей спинного мозга. Пересечение ла терального спинно-таламического пути в вентральных отделах белого вещества спинного мозга приводит к утрате бо левой и температурной чувствительно сти на противоположной стороне с уров ня, находящегося на 1—2 сегмента ниже уровня операции.
Болевые и температурные импуль сы, достигшие таламуса, ощущаются, но не дифференциируются. Только при достижении импульсами коры головно го мозга происходит дифференциация боли.
На рис. 1.19 схематически изобра жена топография путей чувствительно сти: их ход (согласно существующим до настоящего времени представлени ям) от задних корешков к тем областям, где они заканчиваются. Все чувстви тельные «третьи нейроны», связываю щие таламус с корой головного мозга, проходят через заднее бедро внутренней капсулы кзади от пирамидного пути и направляются к зоне восприятия чув ствительных импульсов от всего тела, расположенной в задней центральной (или постцентралыюй) извилине, цитоархитектоническим полям Бродмана За, ЗЬ, 2 и 1. Сюда «третьи нейроны» проводят импульсы поверхностной чув ствительности — болевой, температур ной, тактильной, давления, а также, в некоторой степени, и импульсы глубо кой чувствительности (рис. 1.20 и 55). Не все афферентные импульсы идут из таламуса к чувствительным зонам коры. Часть из них направляется к двигатель
Спинно-таламические пути |
23 |
ным зонам коры, в прецентралъную из вилину. В то же время постцентралыюй извилине могут быть свойственны не только чувствительные, но и двигатель ные реакции. Двигательные и чувстви тельные корковые поля частично пере крываются. Пэтому можно говорить о центральных извилинах как о сенсомоторной области. В этой зоне чувстви тельные сигналы могут быть мгновенно преобразованы в двигательные реакции благодаря наличию сенсомоторной об ратной связи, которая будет рассмотре на ниже. Пирамидные волокна от этих замкнутых кругов обратной связи обыч но идут непосредственно к клеткам пе редних рогов спинного мозга, без во влечения вставочных нейронов. Хотя между пре- и постцентралыюй извили нами и существуют частичные пере крытия связей и функций, но прецентральная извилина до настоящего вре мени считается основной двигательной зоной, а постцентральная — основной чувствительной зоной коры головного мозга. Импульсы, поступающие в кору головного мозга по афферентным во локнам «третьих нейронов», распреде ляются в ней в определенном соматотопическом порядке, корреспондируя со схемой чувствительности человека, сто ящего на голове. Более того, различные виды чувствительности имеют различ ное пространственное представительст во: поле Бродмана За получает импуль сы от мышечных веретен; поле ЗЬ — болевые и температурные импульсы; ноле 1 — тактильные импульсы; поле 2 — импульсы давления (см.. рис. 1.20 и 55).
Как уже отмечалось выше, болевые, температурные и другие импульсы по достижении таламуса воспринимаются в виде неотчетливых, неясных ощуще ний. Дифференциация качества этих импульсов в нашем сознании происхо дит лишь тогда, когда они достигают коры. Более высокоорганизованные ви-
24 1 Система чувствительности
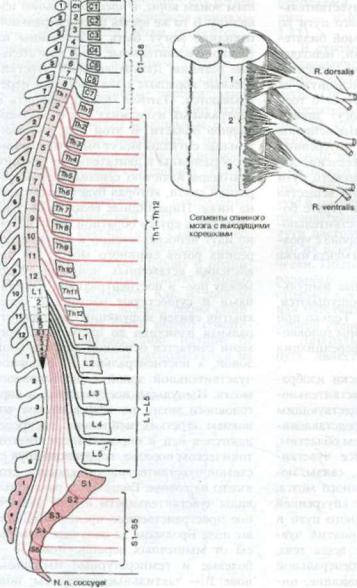
Рис. 1.22. Соотношение между сегментами спинного мозга с выходящими из них корешка ми и телами позвонков.
cl filum tonninalc
ды чувствительности, такие, как дву мерная дискриминационная чувстви тельность, точная индикация локализа ции каждого раздражения в отдельно сти, реализуются в коре головного моз га. Повреждение чувствительных зон коры вызывает лишь уменьшение вос
приятия болевых, температурных и так тильных раздражителей, тогда как дис криминационная чувствительность и чувство позы полностью исчезают в со ответствующих областях противопо ложной поражению стороны тела (по скольку все чувствительные пути преж-
де чем поступить в кору, пересекают среднюю линию и переходят на проти воположную сторону).
Такие функции, как, к примеру, стереогноз (узнавание предметов на ощупь) требуют участия дополнительных ассо циативных зон. Эти зоны располагают ся в теменной доле, где интегрируется множество отдельных ощущений, каса ющихся размера, формы, физических свойств предмета (острота или тупость, мягкость, тяжесть, тепло или холод, и т. д.), и происходит сравнение возник шего образа с тактильными ощущения ми, имевшими место ранее. Поврежде ние нижней теменной доли может при вести к нарушению узнавания предме тов на ощупь на стороне, противопо ложной стороне поражения. Нарушение этой способности называется астерео-
гнозом.
Спинной мозг содержит не только афферентные пути и собственные внут ренние соединительные волокна, такие, как собственные пучки, fasciculi proprii,
Спинно-таламические пути |
25 |
но и ряд эфферентных путей. К ним относятся пирамидные пути, обеспечи вающие выполнение произвольных движений, и, кроме того, многочислен ные так называемые экстрапирамидные пути, оказывающие влияние на слож ные рефлекторные механизмы спинно го мозга. На рис. 1.21 изображен попе речный срез спинного мозга и указано расположение различных чувствитель ных, а также нисходящих двигательных путей и их соотношение друг с другом. Показан ход волокон чувствительных «вторых нейронов». Эти клетки распо ложены в задних рогах, а их аксоны направляются к нейронам ствола мозга
всоставе переднебоковых канатиков.
Кчислу этих путей принадлежат, например, спинно-ретикулЯрный, trac tus spinoreticularis, спинно-тектальный, tractus spinotectalis, спинно-оливарный, tractus spinoolivaris, и спинно-преддвер- ный, tractus spinovestibularis, пути (рис. 1.21). Они относятся к группе аффе рентных путей, соединяющих круги об-
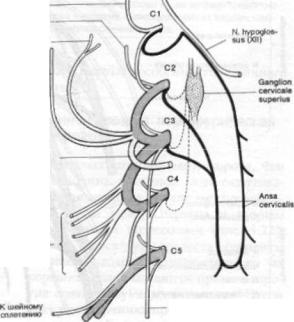
N.suboccipilaliB — N. occipitalis major
N. occipitalis minor
N. auricular is magnus
N. transversus colli
N. n. supradavi culares
N. phrenicus
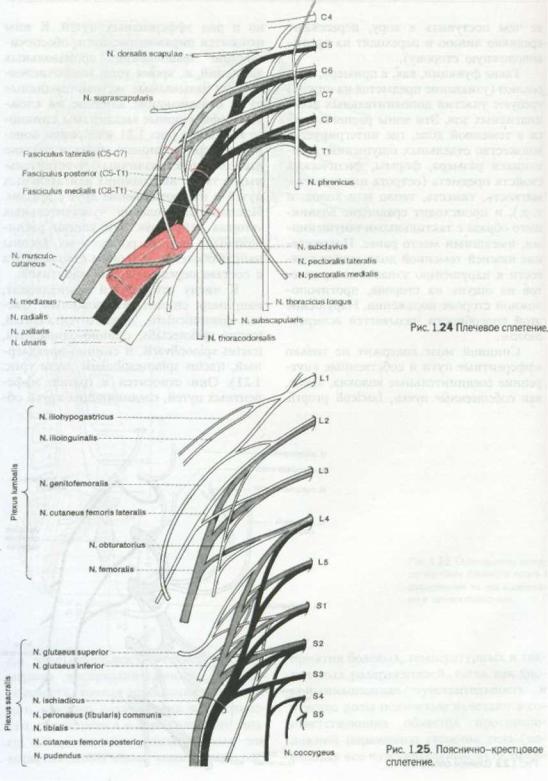
Рис. 1.23. Шейное сплетение.
|
|
|
Спинной мозг и периферическая иннервация |
27 |
|
|
Нврмшй |
|
|
|
|
корешок |
|
|
|
||
Спинной |
; |
Спли.енио |
Периферический нерв |
Дсрмаюм |
|
мозг |
|
|
|||
|
|
|
|
|
|
|
|
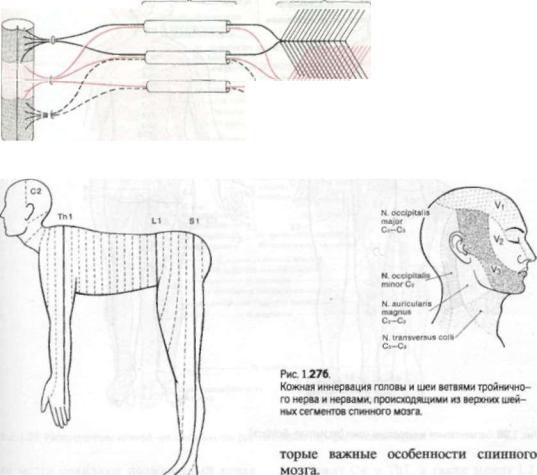
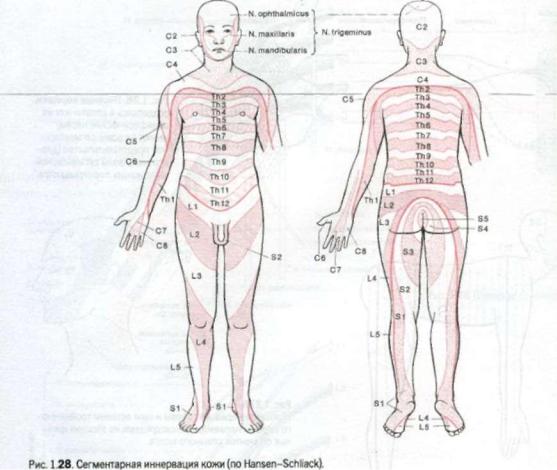
Рис. 126. Нервные корешки, |
' |
/////-////','./' |
разделяясь в сплетениях на |
|
||
W |
//•''/''/'/•'//,' |
перИчЬерические нервы, |
|
|
имеют на коже сегментар |
|
v l l ( ( ( U W U l ( |
ное представительство (дер- |
|
\ \ \ \ \ \ \ \ \ \ \ \ \ \ \ |
матомы). Зоны сегментарной |
|
\ \ \ \ \ \ \\\\\\\\> |
|
|
иннервации перекрываются. |
|
|
|
Рис. 1.27а.
Упрощенная схема сегментарной иннервации
ратной связи экстрапирамидной систе мы, о которых речь пойдет ниже. Спин- но-преддверный путь расположен в шей ном отделе спинного мозга выше уров ня С4 и прилежит к преддверно-спиналь- ному пути, tractus vestibulospinalis. Воз можно, он является коллатералью дор сального спинно-мозжечкового пути.
Теперь необходимо отметить неко-
Спинной мозг и периферическая иннервация
У взрослых спинной мозг короче, чем позвоночник. Спинной мозг оканчива ется примерно на уровне межпозвонко вого диска между первым и вторым поясничными позвонками (рис. 1.22). До 3-х месяного возраста сегменты спинного мозга, обозначаемые по их корешкам, располагаются прямо напро тив соответствующих позвонков. Затем рост позвоночника происходит быстрее,
28 1 Система чувствительности
чем рост спинного мозга. Корешки, на |
ся на уровне третьего поясничного по |
||||||
правляясь к свои межпозвонковым от |
звонка. |
|
|||||
верстиям, идут косо вниз и становятся |
За исключением сегментарного от- |
||||||
все длиннеее и длиннее по направлению |
хождения нервных корешков, в самом |
||||||
к концу спинного мозга (конусу спин |
спинном мозге не обнаруживается мор |
||||||
ного мозга), который обычно находится |
фологических признаков метамерного |
||||||
деления (см. рис. 1.22). Несоответствие |
|||||||
на уровне |
2-го поясничного |
позвонка. |
|||||
между локализацией сегментов спинно |
|||||||
Ниже этого уровня |
субарахноидальное |
||||||
го мозга и соответствующих позвонков, |
|||||||
пространство, |
имеющее здесь вид сум |
||||||
увеличивающееся по мере приближения |
|||||||
ки, содержит |
лишь |
передние |
и задние |
||||
к конусу, должно учитываться |
при оп |
||||||
корешки, |
которые |
образуют |
конский |
||||
ределении уровня поражения |
спинного |
||||||
хвост (рис. |
2.34). В |
редких случаях ко |
|||||
мозга. |
|
||||||
нус спинного мозга может заканчивать |
Корешки CI—С7 сегментов спинно- |
|