

Межполушарные различия при зрительном восприятии
|
Левое полушарие |
Правое полушарие |
|
Лучше узнаются стимулы | |
|
Вербальные |
Невербальные |
|
Легко различимые |
Трудно различимые |
|
Знакомые |
Незнакомые |
|
Лучше решаются задачи | |
|
Оценка временных отношений |
Оценка пространственных отношений |
|
Установление сходства |
Установление различий |
|
Установление идентичности стимулов по названиям |
Установление физической идентичности стимулов |
|
Переход к вербальному кодированию |
Зрительно—пространственный анализ |
|
Особенности процессов восприятия | |
|
Аналитическое восприятие |
Целостное восприятие (гештальт)
|
|
Последовательное восприятие |
Одновременное восприятие |
|
Абстрактное, обобщенное, инвариантное узнавание |
Конкретное узнавание |
|
Предполагаемые морфофизиологические различия | |
|
Фокусированное представительство |
Диффузное представительство |
Выделяют несколько видов функциональных асимметрий. Неодинаковость двигательной активности рук, ног, лица, половин тела, управляемой каждым полушарием мозга, называется моторной асимметрией. Неравнозначность восприятия каждым из полушарий объектов, расположенных слева и справа от средней плоскости тела, именуется сенсорной асимметрией. Специализация полушарии мозга в отношении различных форм психической деятельности обозначается как психическая асимметрия.
Так, человек с превалированием левополушарных функций тяготеет к теории, имеет большой словарный запас и активно им пользуется, ему присуща двигательная активность, целеустремленность, способность прогнозировать события. Правополушарный» человек тяготеет к конкретным видам деятельности, он медлителен и неразговорчив, наделен способностью тонко чувствовать и переживать, он склонен к созерцательности и воспоминаниям.
Совокупность индивидуальных особенностей психики и поведения человека составляет тип высшей нервной деятельности, или темперамент человека ин складывается из общих свойств нервной системы, которые характеризуются экстра—интраверсией, эмоциональной стабильностью — невротизмом, и подвижностью или инертностью нервных процессов.
И. П. Павлов выделял два типа высшей нервной деятельности человека, которые сейчас могут рассматриваться с позиции межполушарной асимметрии—мыслительный тип с преобладанием второй (речевой) сигнальной системы —преимущественно левополушарный и художественный тип с преобладанием первой сигнальной системы конкретных образов — преимущественно правополушарный человек. В последнее время получает признание концепция о взаимодополняющем сотрудничестве двух полушарий и преимуществе отдельного полушария лишь в определенные стадии той или иной нервно—психической деятельности, а не всей функции в целом. Правое полушарие быстрее, чем левое, обрабатывает поступающую информацию, зрительно—пространственный анализ стимулов в правом полушарии передается в левое полушарие (в моторный «центр речи»), где происходит окончательный, высший семантический анализ и осознание раздражения.
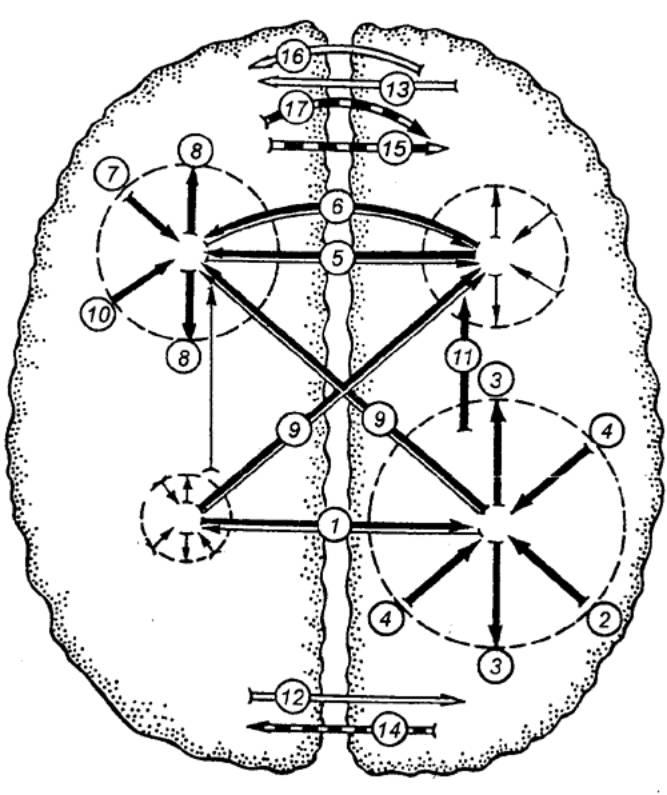
Согласно концепции В. Л. Бианки, в процессе обучения правое полушарие работает по принципу дедукции,т. е. сначала осуществляет синтез, а затем анализ; левое же полушарие функционирует по принципуиндукции,сначала анализируя раздражители, а затем синтезируя их (рис. 3.57).
В эволюции человека речь первоначально лишь использовала, а затем развила и качественно преобразовала уже имеющуюся у животных функциональную межполушарную асимметрию.
|
|
|
Рис. 3.57 Схема межполушарных взаимоотношений (по Бианки) Вверху — ассоциативная кора, внизу — проекционная; слева — левое полушарие; справа — правое; жирные стрелки — доминирующие влияния, тонкие — недоминирующие; белые стрелки — облегчающие влияния; прерывистые — тормозящие; 1,5 — транскаллозальные (через мозолистое тело) влияния; 2,7, 10 — восходящие афферентные влияния; 3, 8 — дивергенция возбуждения; 4 — конвергенция; 6 — экстракаллозальные влияния; 9 — межзональные транскаллозальные влияния; 11 — межзональные внутриполушарные влияния; 12, 13 — транскаллозальные облегчающие влияния; 14, 15 — транскаллозальные тормозящие влияния; 16 — экстракаллозальные облегчающие влияния; 17 — экстракаллозальные тормозящие влияния. Долгое время считалось, что оба полушария мозга абсолютно самостоятельны и представляют собой такой же парный орган, как почки или легкие. Но работы Роджера Уолкотта Сперри по перерезке комиссур мозга показали, что при нарушении взаимосвязи между левым и правым полушариями возникают серьезные нарушения психических функций. Результат перерезки комиссур мозга (мозолистого тела), получивший название «Модель расщепленного мозга», характеризуется триадой симптомов: — аномия, проявляющаяся в нарушении способности давать отчет обо всех видах сенсорной информации, поступающей в правое полушарие; — дископия-дисграфия, для которой характерно нарушение письма и конструктивной деятельности при выполнении их одной (правой или левой) рукой; — нарушение координации движений. Эксперименты показали, что при частичной перерезке мозолистого тела эти симптомы проявляются не полностью. Это связано с тем, что мозолистое тело не является однородным органом, а представляет собой дифференцированную систему, различные участки которого выполняют разные функции в механизмах межполушарного взаимодействия. При частичной перерезке отдельных участков мозолистого тела возникают нарушения только одной модальности. Кроме того, при частичной перерезке мозолистого тела отмечается сравнительно быстрое восстановление психических функций. Работы под руководством Э.Г. Симерницкой показали, что процессы межполушарного взаимодействия у детей протекают иначе, чем у взрослых. У детей синдрома расщепленного мозга не наблюдается, поскольку мозолистое тело созревает достаточно поздно (примерно к двадцати годам). Системный характер психической деятельности состоит в дифференцированном участии различных мозговых образований и полушарий мозга в организации высших психических функций. Ни одно полушарие не может быть рассмотрено как доминирующее по отношению к какой-либо функции или деятельности. Каждое полушарие доминирует по свойственному ему принципу работы, по тому вкладу, которое оно вносит в организацию психической функции. Таким образом, можно сказать, что межполушарная организация психических процессов основана на единстве двух факторов: функциональной асимметрии (специализации) полушарий мозга и их взаимодействии в психической деятельности человека.
|
Речь представляет собой одну из сложных высших психических функций человека.
Речевой акт осуществляется сложной системой органов, в которой главная, ведущая роль принадлежит деятельности головного мозга.
Еще в начале XX в. была распространена точка зрения, по которой функцию речи связывали с существованием в мозгу особых «изолированных речевых центров». И. П. Павлов дал новое направление этому взгляду, доказав, что локализация речевых функций коры головного мозга не только очень сложна, но и изменчива, почему и назвал ее «динамической локализацией».
В настоящее время благодаря исследованиям П. К- Анохина, А. Н. Леонтьева, А. Р. Лурии и других ученых установлено, что основой всякой высшей психической функции являются не отдельные «центры», а сложные функциональные системы, которые расположены в различных областях центральной нервной системы, на различных ее уровнях и объединены между собой единством рабочего действия.
Речь — это особая и наиболее совершенная форма общения, присущая только человеку. В процессе речевого общения (коммуникаций) люди обмениваются мыслями и воздействуют друг на друга. Осуществляется речевое общение посредством языка. Язык — это система фонетических, лексических и грамматических средств общения. Говорящий отбирает необходимые для выражения мысли слова, связывает их по правилам грамматики языка и произносит путем артикуляции речевых органов.
Для того чтобы речь человека была членораздельной и понятной, движения речевых органов должны быть закономерными и точными. Вместе с тем эти движения должны быть автоматическими, т. е. такими, которые осуществлялись бы без специальных произвольных усилий. Так и происходит на самом деле. Обычно говорящий следит только за течением мысли, не задумываясь над тем, какое положение должен занять его язык во рту, когда надо вдохнуть и т. д. Это происходит в результате действия механизма произнесения речи. Для понимания действия механизма произнесения речи необходимо хорошо знать строение речевого аппарата.
Речевой аппарат состоит из двух тесно связанных между собой частей: центрального (или регулирующего) речевого аппарата и периферического (или исполнительного) (рис. 1).
Центральный речевой аппарат находится в головном мозге. Он состоит из коры головного мозга (преимущественно левого полушария), подкорковых узлов, проводящих путей, ядер ствола (прежде всего продолговатого мозга) и нервов, идущих к дыхательным, голосовым и артикуляторным мышцам.
Речь, как и другие проявления высшей нервной деятельности, развивается на основе рефлексов. Речевые рефлексы связаны с деятельностью различных участков мозга. Однако некоторые отделы коры головного мозга имеют главенствующее значение в образовании речи. Это лобная, височная, теменная и затылочная доли преимущественно левого полушария мозга (у левшей правого). Лобные извилины (нижние) являются двигательной областью и участвуют в образовании собственной устной речи (центр Брока). Височные извилины (верхние) являются речеслуховой областью, куда поступают звуковые раздражения (центр Вернике). Благодаря этому осуществляется процесс восприятия чужой речи. Для понимания речи имеет значение теменная доля коры мозга. Затылочная доля является зрительной областью и обеспечивает усвоение письменной речи (восприятие буквенных изображений при чтении и письме). Кроме того, у ребенка речь начинает развиваться благодаря зрительному восприятию им артикуляции взрослых.
Подкорковые ядра ведают ритмом, темпом и выразительностью речи.
Проводящие пути. Кора головного мозга связана с органами речи (периферическими) двумя видами нервных путей: центробежными и центростремительными.
Центробежные (двигательные) нервные пути соединяют кору головного мозга с мышцами, регулирующими деятельность периферического речевого аппарата. Центробежный путь начинается в коре головного мозга в центре Брока.
От периферии к центру, т. е. от области речевых органов к коре головного мозга, идут центростремительные пути.
Центростремительный путь начинается в проприорецепторах и в барорецепторах.
Проприорецепторы находятся внутри мышц, сухожилий и на суставных поверхностях двигающихся органов.
Рис. 1. Строение речевого аппарата: 1 — головной мозг: 2 — носовая полость: 3 — твердое нёбо; 4 — ротовая полость; 5 — губы; 6 — резцы; 7 — кончик языка; 8 — спинка языка; 9 — корень языка; 10 — надгортанник: 11 — глотка; 12 -- гортань; 13 — трахея; 14 — правый бронх; 15 — правое легкое: 16 — диафрагма; 17 — пищевод; 18 — позвоночник; 19 — спинной мозг; 20 — мягкое нёбо
Проприорецепторы возбуждаются под действием мышечных сокращений. Благодаря проприорецепторам контролируется вся наша мышечная деятельность. Барорецепторы возбуждаются при изменениях давления на них и находятся в глотке. Когда мы говорим, происходит раздражение проприои барорецепторов, которое идет по центростремительному пути к коре головного мозга. Центростремительный путь играет роль общего регулятора всей деятельности речевых органов,
В ядрах ствола берут начало черепно-мозговые нервы. Все органы периферического речевого аппарата иннервируются (СНОСКА: Иннервация — обеспеченность какого-либо органа или ткани нервными волокнами, клетками.) черепно-мозговыми нервами. Главные из них: тройничный, лицевой, языкоглоточный, блуждающий, добавочный и подъязычный.
Тройничный нерв иннервирует мышцы, приводящие в движение нижнюю челюсть;лицевой нерв — мимическую мускулатуру, в том числе мышцы, осуществляющие движения губ, надувание и втягивание щек;языкоглоточный иблуждающий нервы — мышцы гортани и голосовых складок, глотки и мягкого нёба. Кроме того, языкоглоточный нерв является чувствительным нервом языка, а блуждающий иннервирует мышцы органов дыхания и сердца.Добавочный нерв иннервирует мышцы шеи, аподъязычный нерв снабжает мышцы языка двигательными нервами и сообщает ему возможность разнообразных движений.
Через эту систему черепно-мозговых нервов передаются нервные импульсы от центрального речевого аппарата к периферическому. Нервные импульсы приводят в движение речевые органы.
Но этот путь от центрального речевого аппарата к периферическому составляет только одну часть речевого механизма. Другая его часть заключается в обратной связи — от периферии к центру.
Нарушение речи возникает вследствие органического поражения головного мозга, тех его участков мозга, которые отвечают за речевую зону, происходит искаженное произношение звуков. Причины органического поражения вызываются нарушением мозгового кровообращения, опухолями головного мозга, пороками развития нервной системы.
Психопатологические симптомынарушения или распада речи зависят от локализации ОНМК. Нарушения речи проявляется в виде полной или частичной утраты речи (тотальная и моторная афазия); нарушения звукопроизносительной стороны речи (дизартрия) и нарушения темпа речи в виде спотыкания.
Поражение разных участков коры головного мозга приводит к качественно отличным нарушениям речи. В связи с этим, выделяются различные области, поражение которых вызывает те или иные расстройства речи.
моторный , так называемый «Центр Брока» производит речь, регулируя голосовой аппарат (голосовые связки, мышцы гортани), управляя речевой и дыхательной мускулатурой и всем тем, что участвует в произношении звуков. Именно, в «зоне Брока», расположенной в задней части нижней лобной извилины левого полушария (у правшей), — центре моторной речи — формируется двигательная программа речевого высказывания.
сенсорный или чувствующий «Центр Вернике» — слуховой центр речи, обеспечивает способность слышать, распознавать и контролировать своюи чужую речь. «Зона Вернике» — центр восприятия речи — расположена в заднем отделе верхней височной извилины левого полушария (у правшей).
при помощи третьих «ассоциативных» (объединяющих) центров, расположенных в нижней теменной доле и отвечающих за правильный подбор слов, мы анализируем, «думаем», что говорить дальше, создавая связную структуру фраз и выстраивая предложения.
Дизартрия- Одна из форм расстройства экспрессивной речи, обусловленная нарушением функций исполнительного речевого аппарата (вялый или спастический парез/паралич, ригидность, атаксия, заикание). При полной сохранности понимания речи окружающих, письма и чтения про себя нарушается собственная артикулированная речь; словарь и грамматический строй ее при этом остаются ненарушенными. Особенно страдает артикуляция согласных, речь замедлена, иногда прерывиста. Является, в частности, одним из проявлений бульбарного синдрома и псевдобульбарного синдрома, следствием поражения мозжечка и его связей, базальных узлов, премоторных отделов коры доминантного полушария.
Афазия— это расстройство речи, состоящее в утрате способности пользоваться словами и фразами для выражения мысли.При афазии частично или полностью утрачивается возможность пользоваться словами для выражения мыслей и общения с окружающими при сохранности функций артикуляционного аппарата (органы, отвечающие за произношение звуков) и слуха. Афазия может носить стойкий или временный характер. Временные нарушения речи обусловлены, прежде всего временными нарушениями кровообращения в головном мозге, возникающими в результате сужения артерий, питающих головной мозг. Иногда временные нарушения речи развиваются при некоторых разновидностях мигрени. Стойкая афазия возникает при серьезном поражении головного мозга. Афазия подразделяется на моторную, сенсорную и амнестическую.Моторная афазия— характеризуется утратой способности к произношению слов, речь становится маловыразительной, замедленной, произносимые слова искажены в результате затруднения формирования звуков и слогов, а также целых слов. Страдает также процесс переключения с одной речевой единицы (звук, слова) на другую. Внешне это проявляется так называемой персеверацией - застреванием на одном слоге или слове. Грамматическое строение речи неправильное (аграмматизм). Симптомы моторной афазии: в особо тяжелых случаях речь невозможна, а в более легких затруднена. Моторная афазия в чистом виде встречается редко, обычно расстройства речи сочитаются с нарушением письма (аграфия). Мотроная афазия проявляется при возникновении патологического очага в задней трети нижней лобной извилины (центр Брока).Сенсорная афазия- утрата способности понимания речи окружающих, вплоть до отсутствия реакции на нее в тяжелых случаях. Хотя бывает, что элементарный слух сохранен, но разобрать речь, разделить ее на отдельные слова больной не может. Вследствие отсутствия слухового восприятия нарушается и собственно речь. Больные сенсорной афазией говорят много, но произносимые слова непонятны, поскольку один звук (буква) заменяется другим. В результате речь представляет собой набор бессмысленных звуков. У больного отсутствует сознание своего речевого дефекта. Обычно сенсорная афазия сочетается с нарушением понимания письма (алексия). При сенсорной афазии потологический очак локализуется в задних отделах верхней височной извилины (центр Вернике). Сочетание моторной и сенсорной афазий обозначается кактотальная афазия. Проявляется тотальная афазия, как правило при обширных очагах воспаления или кровоизлияния в головной мозг.
Амнестическая афазия - проявляется забыванием называния предметов. Выпадения из памяти названия предметов является наиболее характерной чертой данного вида афазии. Обычно небольшая подсказка, например первых букв, слогов, помогает больному, но чаще качестве замены больной использует обозначение функции или качества предмета. Амнестическая афазия редко бывает в чистом виде, чаще наблюдается с сенсорной афазией. Локализация очага на стыке височно-теменно-затылочной области.
Механизмы целенаправленной деятельности человека. Физиология эмоций.
Представление о механизме целенаправленной деятельности развивалось поэтапно. По Павлову узловым механизмом ВНД является условный рефлекс. Однако поведение человека в конкретных условиях нельзя объяснить суммой условно-рефлекторных реакций. Поведенческая деятельность организма - это не только набор реакций, обеспечивающих его приспособление к условиям среды, но и активные действия человека, направленные не изменение этих условий. Павлов ввел понятие системности в работе коры больших полушарий, понимая под этим динамическое объединение различных структур мозга для осуществления конкретных целей. Организм человека И.П.Павлов рассматривал как «систему в высшей степени саморегулирующуюся – саму себя поддерживающую».
Позже А.А. Ухтомский установил, что ответы животных на внешние раздражители существенно зависят от формирующихся в организме доминантных состояний.
Теория условных рефлексов и теория доминантности явились закономерной предпосылкой возникновения теории функциональных систем П.К. Анохина.
Теория функциональных систем исходит из того, что единицами целостной деятельности организма являются динамические, самоорганизующиеся системы, компоненты которой взаимодействуют между собой с целью достижения полезного приспособительного результата, например, удовлетворение потребности в чем-то.
В теории функциональных систем П.К. Анохин предпринял попытку объяснить любую поведенческую целенаправленную деятельность организма на основе алгоритма.
Функциональные системы возникают на основе нескольких источников: показателей внутренней среды и результатов поведенческой деятельности, удовлетворяющих биологические потребности.
При формировании функциональных систем закладывается прогноз: а) какой результат должен быть получен в итоге того или иного поведения; б) с помощью каких механизмов должен быть получен результат; в) как оценить достижение результата.
В соответствии с представлениями П.К. Анохина любая целенаправленная деятельность в своем развитии включает последовательно сменяющиеся стадии:
а) афферентный синтез;
б) принятие решения;
в) формирование акцептора результата действия;
г) эфферентный синтез;
д) совершаемое действие;
е) оценка результата действия.
В развитии любой из названных стадий участвуют определенные структуры мозга.
Афферентный синтез необходим для принятия решения: какое поведение является определяющим в данных условиях. Он формируется на основании доминирующей мотивации, в основе которой лежит актуальная потребность; с учетом имеющегося опыта (памяти о том, как поступить в подобных условиях). При этом в ЦНС происходит анализ обстановочной и пусковой афферентации – комплекса раздражителей, оценивая которые, человек решает возможно ли в этих условиях выбранное поведение.
Из структур ЦНС в афферентном синтезе участвуют: гипоталамус и миндалевидный комплекс - они обеспечивают формирование доминирующей мотивации; ретикулярная формация; центры памяти; лимбическая система и сенсорная кора больших полушарий.
Принятие решения сопровождается формированием программы действия, т.е. набора команд к исполнительным органам.
Важнейшее значение в принятии решения имеют лобные доли коры больших полушарий.
Формирование акцептора результата действия – это создание нейронной матрицы или копии предполагаемого результата действия, если будет выполнено принятое решение.
Эфферентный синтез обеспечивает формирование определенных команд, которые поступят к исполнительным органам – эффекторам (скелетным мышцам, внутренним органам, железам). Его задача выбрать оптимальную для данных условий программу действий.
Совершается действие, которое сопровождается определенным результатом. Информация о реально достигнутом результате по каналам обратной связи поступает в блок акцептора результата действия, сравнивается с копией предполагаемого результата. В случае, если полученный результат соответствует ожидаемому, функциональная система выполнила свою задачу и прекращает свое действие. Если результат не достигнут, формируется новая функциональная система с новой программой действий.
В этом и заключается динамичность функциональной системы.
Важно отметить, что с позиций функциональной системы нельзя объяснить проявление высших психических функций: озарение, творчество, сознание и др.
Таким образом, функциональная система представляет временное объединение различных элементов нервной системы; формируется на основе доминирующей потребности и обеспечивает выполнение конкретной задачи: совершить действие для удовлетворения возникшей потребности.
Эмоции– это реакции, отражающие субъективное отношение индивида к объективным явлениям. Эмоции возникают в составе мотивации и играют важную роль в формировании поведения. Выделяют 3 вида эмоциональных состояний (А. Н. Леонтьев):
1. Аффекты – сильные, кратковременные эмоции, возникающие на уже имеющуюся ситуацию. Страх, ужас при непосредственной угрозе жизни.
2. Собственно эмоции – длительное состояние, отражающее отношение индивида к изменившейся или ожидаемой ситуации. Печаль, тревога, радость.
3. Предметные чувства – постоянные эмоции, связанные с каким-либо объектом (чувство любви к конкретному человеку и т.д.).
Функции эмоций:
1. Оценочная. Она позволяет быстро оценить возникающую потребность и возможность ее удовлетворения. Например, при чувстве голода человек не подсчитывает калорийность имеющейся пищи, содержание в ней белков, жиров, углеводов, а просто ест в соответствии с интенсивностью чувства голода, т.е. интенсивностью соответствующей эмоции.
2. Побуждающая. Эмоции стимулируют целенаправленное поведение. Например, отрицательные эмоции при голоде стимулируют пищедобывающее поведение.
3. Подкрепляющая. Эмоции стимулируют запоминание и обучение. Например, положительные эмоции при материальном подкреплении обучения.
4. Коммуникативная. Состоит в передаче своих переживаний другим индивидам. Эмоции передаются с помощью мимики, а не мысли.
Эмоции выражаются определенными двигательными и вегетативными реакциями. Например, при определенных эмоциях возникает соответствующая мимика, жестикуляция. Возрастает тонус скелетных мышц. Изменяется голос. Учащается сердцебиение, повышается артериальное давление. Это объясняется возбуждением двигательных центров, центров симпатической нервной системы и выбросом адреналина из надпочечников (полиграфия).
Основное значение в формировании эмоций принадлежит гипоталамусу и лимбической системе. Особенно миндалевидному ядру. При его удалении у животных механизмы эмоций нарушаются. При раздражении миндалевидного ядра у человека страх, ярость, гнев.
У человека важное значение в формировании эмоций принадлежит лобной и височной областям коры. Например, при повреждении лобных областей возникает эмоциональная тупость. Неодинаково и значение полушарий. При временном выключении левого полушария возникают отрицательные эмоции – настроение становится пессимистичным. При выключении правого – возникает противоположное настроение.
Установлено, что первоначально чувство благодушия, беспечности, легкости при употреблении алкоголя объясняется его воздействием на правое полушарие. Последующее ухудшение настроения, агрессивность, раздражительность обусловлено действием алкоголя на левое полушарие. Поэтому у людей с недостаточно развитым левым полушарием алкоголь практически сразу вызывает агрессивное поведение. У здоровых людей эмоциональное преобладание правого полушария проявляется мнительностью, повышенной тревожностью. При доминантности левого – этих явлений нет (тест эмоциональной асимметрии мозга - юмор).
Важное значение в возникновении эмоций принадлежит балансу нейромедиаторов. Например, если в мозге возрастает содержание серотонина, настроение улучшается, при его недостатке наблюдаются депрессии. Такая же картина наблюдается при недостатке или избытке норадреналина. Обнаружено, что у самоубийц значительно снижено содержание этих нейромедиаторов в мозге.
Учение И. П. Павлова об анализаторах. Структура анализатора.
СЕНСОРНАЯ СИСТЕМА– это совокупность специализированных нервных образований, обеспечивающих кодирование и декодирование физических характеристик сенсорных сигналов.
Учение об анализаторах было создано И.П.Павловым, который рассматривал АНАЛИЗАТОРкак единую систему, включающую ТРИ ОТДЕЛА, функционально и анатомически связанных друг с другом:
периферическийили рецепторный(включает рецепторный аппарат);
проводниковый(представлен афферентным и промежуточными нейронами);
центральныйиликорковый(представлен участками коры больших полушарий, воспринимающими афферентные сигналы).
Общие принципы строения и функционирования анализаторов.
Сенсорная система включает следующие элементы:
вспомогательный аппарат;
сенсорный рецептор;
сенсорные пути;
проекционная зона коры больших полушарий.
Вспомогательный аппарат представляет собой образование, функцией которого является первичное преобразование энергии действующего стимула.
Например, вспомогательный аппарат вестибулярной системы преобразует угловые ускорения тела в механическое смещение киноцилей волосковых клеток.
Вспомогательный аппарат характерен не для всех сенсорных систем.
Сенсорный рецептор осуществляет преобразование энергии действующего раздражителя в специфическую энергию нервной системы, то есть в упорядоченную последовательность нервных импульсов.
В первичном рецепторе эта трансформация осуществляется в окончаниях чувствительного нейрона, а во вторичном рецепторе она происходит в рецепторной клетке.
Аксон чувствительного нейрона (первичный афферент) проводит нервные импульсы в ЦНС.
В ЦНС возбуждение передается по цепочке нейронов (так называемый сенсорный путь) к коре больших полушарий.
Аксон чувствительного (сенсорного) нейрона образует синаптические контакты с несколькими вторичными сенсорными нейронами. Аксоны последних следуют к нейронам, расположенным в ядрах более высоких уровней.
По ходу сенсорных путей происходит обработка информации, в основе которой лежит интегративная деятельность нейрона. Окончательная обработка сенсорной информации происходит в коре больших полушарий.
Основные функции анализаторов.
Рецепция раздражителей и преобразование, то есть трансформация рецепторного сигнала.
Кодирование информации и передача её в виде двоичного кода, то есть потенциала действия, к сенсорным ядрам ЦНС.
Анализ, идентификация свойств и опознание сигнала.
Физиология рецепторов. Классификация рецепторов. Адекватные и неадекватные раздражители рецепторов. Механизм возбуждения рецепторов. Рецепторный или генераторный потенциал. Кодирование в рецепторах.
Рецепторы– специализированные чувствительные образования, воспринимающие раздражения из внешней и внутренней среды организма и преобразующие их в специфическую активность нервной системы.
Клеточные рецепторы– расположены в наружныхи внутреннихмембранах различных клеток организма, осуществляют их взаимодействиес окружающей средой и обмен информациеймежду органеллами внутри клетки (рецепторы мембран ядра, митохондрий, рибосом, лизосом и др.)
Сенсорные рецепторы–высокоспециализированные образования, воспринимающие раздражители внешней и внутренней среды, трансформирующие энергию раздражителя в энергию нервных импульсов (в виде рецепторного потенциала) и кодирующие свойства раздражителя.
Классификация рецепторов:
1. По локализации в организме:
экстерорецепторы – информируют организм об изменениях во внешней среде (слуховые, зрительные, обонятельные, вкусовые, тактильные и др.);
интерорецепторы информируют об изменениях во внутренней среде (вестибулорецепторы, проприорецепторы, хеморецепторы и др.).
2. По модальности (природе) адекватного раздражителя:
механо-, фото-, хемо-, баро-, ноци- и др. рецепторы.
3. По восприятию раздражителей одной или нескольких модальностей:
мономодальные – приспособлены к восприятию раздражителей одной модальности (большинство экстерорецепторов и часть интерорецепторов, например, хеморецепторы каротидной зоны);
полимодальные – приспособлены к восприятию раздражителей нескольких модальностей (большинство интерорецепторов).
4. По скорости адаптации:
быстроадаптирующиеся –зрительные, слуховые, вкусовые;
медленноадаптирующиеся –проприо-, рецепторы растяжения лёгких, др.
5. По структурно-функциональным особенностям:
свободные нервные окончания – тонкие, безмиелиновые терминали дендритов чувствительных нейронов, свободно контактирующие с раздражителем (например, рецепторы прикосновения).
Свойства:
Являются полимодальными.
Имеют самую низкую чувствительностьсреди всех рецепторов.
Адекватный раздражитель– это сигнал, к воздействию которого в процессе эволюции приспособились рецепторы и структуры сенсорной коры.
Например:
звук– для рецепторов уха и слуховой зоны коры;
свет– для рецепторов глаза и зрительной зоны коры.
Критерием адекватностиявляется порог ощущения, который для адекватного сигнала значительно ниже.
Неадекватный раздражитель– это сигнал, который действует на структуру, специальноне приспособленную для его восприятия
Например:
возбуждение фоторецепторов под влиянием механического удара, а не под воздействием света.
Адекватность или неадекватность раздражителя определяется не собственными его качествами, а специфичностью рецепторного аппарата – местоположением рецептора, а также присутствием в нём специфических клеточных органелл определяют особые свойства рецепторов. Для зрительного рецептора адекватным раздражителем является свет. В то же время свет — неадекватный раздражитель для слухового или тактильного рецептора. Только по отношению к адекватному раздражителю рецептор является высоковозбудимым образованием, реагирующим на ничтожную силу раздражителя. Благодаря этому свойству уже на входе сенсорной системы отфильтровываются все раздражения, к восприятию которых данный рецептор не приспособлен. Реакция возникает только на специфическое раздражение, которое и формирует ощущение строго определенного качества, например, зрительное, вкусовое и т.д. Чем выше специфичность сенсорного органа, тем больше вероятность его возбуждения (при физиологических условиях) только адекватными стимулами. Так, для получения эффекта возбуждения в рецепторе глаза требуется несколько квантов света. Сила неадекватного раздражителя, способная вызвать специфический эффект возбуждения, должна быть в десятки и сотни тысяч раз больше. Например, ощущение света («искры из глаз») может вызвать и механический раздражитель. Но он должен превышать силу адекватного раздражителя в миллионы раз.
В случае первичных рецепторов, действие раздражителя воспринимается окончанием чувствительного нейрона. Действующий раздражитель может вызывать гиперполяризацию или деполяризацию поверхностной мембраны рецепторы в основном за счет изменения натриевой проницаемости. Повышение проницаемости к ионам натрия приводит к деполяризации мембраны и на мембране рецептора возникает рецепторный потенциал. Он существует до тех пор, пока действует раздражитель.
Рецепторный потенциалне подчиняется закону «Все или ничего», его амплитуда зависит от силы раздражителя. У него нет периода рефрактерности. Это позволяет суммироваться рецепторным потенциалам при действии последующих раздражителей. Он распространяется мелено, с угасанием. Когда рецепторный потенциал достигает критической пороговой величины, он вызывает появление потенциала действия в ближайшем перехвате Ранвье. В перехвате Ранвье возникает потенциал действия, который подчиняется закону «Все или ничего» Этот потенциал будет распространяющимся.
Во вторичном рецепторе действие раздражителя воспринимается рецепторной клеткой. В этой клетке возникает рецепторный потенциал, следствием которого будет являться выделение медиатора из клетки в синапс, который действует на постсинаптическую мембрану чувствительного волокна и взаимодействие медиатора с рецепторами приводит к образованию другого, локального потенциала, который называют генераторным. Он по своим свойства идентичен рецепторным. Его амплитуда определяется количеством выделившегося медиатора. Медиаторы – ацетилхолин, глутамат.
Потенциалы действия возникают периодически, т.к. для них характерен период рефрактерности, когда мембрана утрачивает свойство возбудимости. Потенциалы действия возникают дискретно и рецептор в сенсорной системе работает, как аналогово-дискретный преобразователь. В рецепторах наблюдается приспособление – адаптация к действию раздражителей. Есть быстроадаптирующиеся, есть медленно адаптирующиеся. При адаптация снижается амплитуда рецепторного потенциала и число нервных импульсов, которые идут по чувствительному волокну. Рецепторы кодируют информацию. Оно возможно по частоте потенциалов, по группировки импульсов в отдельные залпы и интервалами между залпами. Кодирование возможно по числу активированных рецепторов в рецептивном поле.
Амплитуда и длительность отдельных нервных импульсов, поступающих от рецепторов к центрам, при разных раздражениях остаются постоянными. Однако рецепторы передают в нервные центры адекватную информацию не только о характере, но и о силе действующего раздражителя. Информация об изменениях интенсивности раздражителя кодируется двумя способами:
1) изменением частоты импульсов, идущих по каждому из нервных волокон от рецепторов к нервным центрам, и 2) изменением числа и распределения импульсов — их количества в пачке, интервалов между пачками, продолжительности отдельных пачек импульсов, числа одновременно возбужденных рецепторов и соответствующих нервных волокон.
Чем больше интенсивность раздражителя, тем больше частота афферентных нервных импульсов и их количество. Это обусловливается тем, что нарастание силы раздражителя приводит к увеличению деполяризации мембраны рецептора, что, в свою очередь, вызывает увеличение амплитуды генераторного потенциала и повышение частоты возникающих в нервном волокне импульсов. Между логарифмом силы раздражения и числом нервных импульсов существует прямо, пропорциональная зависимость.
Имеется еще одна возможность кодирования сенсорной информации. Избирательная чувствительность рецепторов к адекватным раздражителям уже позволяет отделить различные виды действующей на организм энергии. Однако и в пределах одной сенсорной системы может быть различная чувствительность отдельных рецепторов к разным по характеристикам раздражителям одной и той же модальности.
Различение сигналов. Абсолютные и дифференциальные пороги ощущения. Адаптация анализаторов.
Важная характеристика сенсорной системы – способность замечать различия в свойствах одновременно или последовательно действующих раздражителей. Различение начинается в рецепторах, но в этом процессе участвуют нейроны всей сенсорной системы. Оно характеризует то минимальное различие между стимулами, которое сенсорная система может заметить (дифференциальный, или разностный, порог).
Для того чтобы в результате действия раздражителя на органы чувств возникло ощущение, необходимо, чтобы вызывающий его стимул достиг определенной величины или порога чувствительности. Выделяют два типа порогов чувствительности: абсолютныйидифференциальный(или порог чувствительности к различению).
Абсолютный порог ощущения – наименьший по интенсивности стимул, способный вызвать определённое ощущение. Величина абсолютного порога зависит от:
характеристик действующего стимула (например, абсолютный порог для звуков разной частоты будет различным);
условий, в которых проводится измерение;
функционального состояния организма: направленности внимания, степени утомления и т.п.
Дифференциальный порог ощущения – минимальная величина, на которую один стимул должен отличаться от другого, чтобы эта разница ощущалась человеком.
В 1834 г. Вебер показал, что для различения веса двух предметов их разница должна быть больше, если оба предмета тяжёлые и меньше, если оба предмета лёгкие.
Согласно закону Вебера, величина дифференциального порога ощущения прямо пропорциональна силе действующего стимула.
Адаптация является общим свойством всех анализаторов. Адаптация — это приспособление рецепторов и анализаторов к силе раздражителя. Возбуждение рецептора наиболее интенсивно в первые моменты его раздражения, затем оно резко снижается и при этом не обнаруживается утомления.
Адаптация зависит не только от продолжительности раздражения, но и от силы раздражителя: чем сильнее раздражитель, тем быстрее наступает адаптация. Адаптация обуславливается главным образом физиологическими процессами в мозговых областях анализаторов, а также процессами, совершающимися в самих рецепторах. В основе адаптации лежит колебания лабильности рецепторов и нейронов головного мозга.
Предполагается, что адаптация обусловлена распадом медиатора, происходящего сравнительно быстро в рецепторах и значительно медленнее в мозговых центрах.
Физиологическое значение адаптация во всех анализаторах заключается в установлении оптимального количества сигналов, поступающих в центральную нервную систему. Адаптация характеризуется известной степенью специализации. Так, при адаптации к сильному запаху сохраняется достаточно высокая чувствительность к действию вновь поступившего слабого запаха.
В некоторых случаях в результате адаптации раздражители полностью перестают восприниматься. Например, запахи почвы, грунтовых вод или фоновые запахи на местности в результате длительного пребывания собаки перестают вызывать ощущения по причине адаптации.
Под сенсибилизацией чувствительности анализаторов понимают повышение их возбудимости под влиянием многократных раздражителей. Свойство сенсибилизации проявляется у служебных собак в процессе систематической тренировки анализаторов при работе по запаховым следам, выборке вещей, человека и т.д.
Передача и преобразование сигналов. Кодирование информации. Детектирование и опознание образов.
Субъективное ощущение, возникающее в результате действия сенсорного стимула, обладает рядом характеристик, то есть позволяет определить ряд параметров действующего раздражителя:
качество (модальность);
интенсивность;
временные характеристики (момент начала и окончания действия раздражителя, динамику силы раздражителя);
пространственную локализацию.
Кодирование качества раздражителя в ЦНС основано на принципе специфичности сенсорных систем и принципе соматотопической проекции. Любая последовательность нервных импульсов, возникших в проводящих путях и корковых проекционных зонах зрительной сенсорной системы, будет вызывать зрительные ощущения.
Кодирование интенсивности. С увеличением амплитуды деформации кожи частота импульсации рецепторов (дисков Меркеля или окончаний Руффини) растёт, следовательно, они регистрируют силу или давление, то есть служат рецепторами интенсивности.
Кодирование временных характеристик невозможно отделить от кодирования интенсивности. При изменении во времени силы действующего стимула, будет изменяться и частота потенциалов действия, образующихся в рецепторе. При длительном действии раздражителя постоянной силы частота потенциалов действия постепенно снижается, поэтому генерация нервных импульсов может прекращаться ещё до прекращения действия раздражителя.
Кодирование пространственной локализации. Организм может достаточно точно определять локализацию многих раздражителей в пространстве. Механизм определения пространственной локализации раздражителей основывается на принципе соматотопической организации сенсорных путей.
Обнаружение и первичное различение сигналов обеспечивается рецепторами, а детектирование и опознание сигналов — нейронами коры больших полушарий. Передачу, преобразование и кодирование сигналов осуществляют нейроны всех слоев сенсорных систем.
Зрительный анализатор. Оптическая система глаза. Аккомодация. Аномалии рефракции.
ЗРИТЕЛЬНЫЙ АНАЛИЗАТОР – это совокупность защитных, оптических, рецепторных и нервных структур, воспринимающих и анализирующих световые раздражители.
Световые раздражители представляют собой электромагнитное излучение с различными длинами волн – от длинных (красная часть спектра) до коротких (синяя часть спектра) и характеризуются:
частотой (определяет окраску цвета)
интенсивностью (яркость).
Зрительный анализатор обеспечивает получение более 80 % информации о внешнем мире за счёт:
пространственной разрешающей способности (острота зрения);
временной разрешающей способности (время суммации и критическая частота мельканий);
порога чувствительности, адаптации, способности к восприятию цветов, стереоскопии (восприятие глубины и объёма).
ОРГАН ЗРЕНИЯ включает в себя:
ОПТИЧЕСКУЮ систему глаза;
РЕЦЕПТОРНЫЙ аппарат сетчатки.
Оптическая система включает в себя радужную оболочку, роговицу, глазные среды и хрусталик.
РАДУЖНАЯ ОБОЛОЧКА – определяет количество попадающего в глаз света (парасимпатические влияния суживают, а симпатические - расширяют зрачок).
РОГОВИЦА, ГЛАЗНЫЕ СРЕДЫ и ХРУСТАЛИК образуют эффективную систему фокусировки, создающую изображение на светочувствительной сетчатке.
ХОД ЛУЧЕЙ через оптическую систему глаза определяется:
радиусом преломляющих поверхностей;
показателем преломления сред глаза.
Преломляющая СИЛА тем больше, чем короче ФОКУСНОЕ РАСТОЯНИЕ (растояние от оптического центра системы до той точки, в которой сходятся преломленные лучи);
Приспособление глаза к чёткому видению различно удалённых предметов или фокусирование глаза осуществляется при помощи механизмов АККОМОДАЦИИ, которые обеспечиваются нейрональными элементами подкорковых и корковых зрительных центров, чувствительных к чёткости контуров изображения и регулируются за счёт изменения тонуса ЦИЛЛИАРНОЙ мышцы.
При рассмотрении ДАЛЁКИХ предметов ресничная мышца расслаблена, циннова связка натянута, в результате чего происходит сдавливание (спереди назад) и растягивание хрусталика. Поэтому ЛУЧИ света ФОКУСИРУЮТСЯ на СЕТЧАТКУ.
При рассмотрении БЛИЗКИХ предметов происходят обратные процессы.
В нормальном глазе (ЭММЕТРОПИЧЕСКИЙ глаз) при полностью расслабленной аккомодации изображение достаточно удалённых предметов фокусируется на сетчатке, что обеспечивает их чёткое видение.
Аномалии рефракции:
МИОПИЯ (близорукость) – возникает в удлинённом глазе, когда главный фокус располагается перед сетчаткой.
ГИПЕРМЕТРОПИЯ (дальнозоркость) – возникает в коротком глазе. При этом зона чёткого изображения располагается за сетчаткой.
СФЕРИЧЕСКАЯ АБЕРРАЦИЯ – возникает, когда лучи, проходящие через периферическую часть хрусталика, преломляются сильнее. Следствием является искажение изображения.
ХРОМАТИЧЕСКАЯ АБЕРРАЦИЯ – возникает, когда хрусталик неодинаково преломляет свет различной длины.
АСТИГМАТИЗМ – дефект светопреломляющих сред глаз, связанный с неодинаковой кривизной их преломляющих поверхностей.
ПРЕСБИОПИЯ (старческая дальнозоркость) – возникает в результате постепенной утраты (в течение жизни) хрусталиком своих основных свойств (прозрачности и эластичности). При этом сила аккомодации уменьшается, и точка ближнего ясного видения отодвигается вдаль.
КАТАРАКТА – это помутнение и потеря эластичности хрусталика в результате дегенерации его внутренних слоёв, которые находятся (с точки зрения обмена веществ) в наименее благоприятных условиях.
Восприятие и обработка сигналов в сетчатке.
Рецепторная система зрительного анализатора представлена в СЕТЧАТКЕ, где происходит первичная обработка зрительной информации и преобразование оптических сигналов в биоэлектрические реакции.
Сетчатка имеет многослойное строение и содержит ФОТОРЕЦЕПТОРЫ (включающие палочки и колбочки, которые обеспечивают синтез зрительных пигментов и поглощение световых лучей) и несколько слоев нейронов (передающих рецепторный потенциал на волокна зрительного нерва).
ФОТОХИМИЧЕСКИЙ ПРОЦЕСС ЗРИТЕЛЬНЫХ ПИГМЕНТОВ запускается поглощением одного кванта света одной молекулой пигмента ПАЛОЧКИ (120 млн.) – содержат зрительный пигмент РОДОПСИН и обеспечивают НОЧНОЕ зрение.
КОЛБОЧКИ (6 млн.) – содержат зрительный пигмент ЙОДОПСИН. Они обеспечивают ДНЕВНОЕ зрение и восприятие ЦВЕТА.
В результате распада пигментов (родопсина в палочках и йодопсина в колбочках) через ряд химических превращений образуются белок ОПСИН и витамин А.
ВОССТАНОВЛЕНИЕ (РЕСИНТЕЗ) ПИГМЕНТОВ происходит в темноте в результате цепи химических реакций, протекающих с поглощением энергии с обязательным участием цис-изомера витамина А.
ПРИ ПОСТОЯННОМ ОСВЕЩЕНИИ фотохимический распад пигментов уравновешен с ресинтезом пигментов.
НЕРВНАЯ ПЕРЕДАЧА в СЕТЧАТКЕ: световые лучи проходят все слои сетчатки и поглощаются в наружных сегментах рецепторных клеток, в результате чего запускается фотохимический процесс зрительных пигментов. В результате формируется рецепторный потенциал (гиперполяризационный) в фоторецепторах, который приводит к генерации потенциала действия в биполярных клетках и волокнах зрительного нерва.
Проводниковый и корковый отделы зрительного анализатора.
ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ В НЕРВНЫЕ ЦЕНТРЫ осуществляется по зрительному нерву в продолговатый мозг (мигательный защитный рефлекс).
В передних буграх четверохолмия среднего мозга находятся первичные зрительные центры, которые обеспечивают зрительные ориентировочные рефлексы, рефлекторные движения глаз, зрачковый рефлекс, аккомодацию глаз, сведение зрительных осей.
В задней доле мозжечка находятся центры, отвечающие за движения глаз.
В зрительных буграх гипоталамуса находятся ядра, отвечающие за расширение (задние ядра) зрачков и глазных щелей и сужение (передние ядра) зрачков и глазных щелей.
В таламусе (латеральное коленчатое тело) находится переключающее ядро зрительных сигналов.
В затылочной доле коры головного мозга (17, 18, 19 поля) находится проекционная зона зрительного анализатора, где осуществляется проекция сетчатки глаз.
В лобной и теменной долях коры головного мозга находится ассоциативная зона зрительного анализатора.
Восприятие цвета, световая и темновая адаптация, восприятие пространства.
В основе восприятия цвета лежат сложные физико-химические процессы, совершающиеся в зрительных рецепторах. Различают три типа «колбочек», проявляющих наибольшую чувствительность к трем основным цветам видимого спектра:
красно-оранжевому (600 – 700 нм);
зеленому (500 – 600 нм);
синему (400 – 500 нм).
Особенности цветовой чувствительности клеток определяются различиями в зрительном пигменте. Комбинации возбуждений этих приемников разных цветов дают ощущения всей гаммы цветовых оттенков.
В компьютерной промышленности эти цвета называются тремя первичными цветами – RGB (Red, Green, Blue). Все цвета, встречающиеся в природе, можно создать, смешивая свет трех этих длин волн и варьируя их интенсивность. Смесь, состоящая из 100% каждого цвета, дает белый свет. Отсутствие всех цветов дает отсутствие света или черный свет.
В случае ослабления восприятия одного из цветов цветовое зрение может нарушаться. Известны три разновидности частичной цветовой аномалии: «краснослепые», «фиолетослепые» и «зеленослепые». Впервые нарушение цветового зрения было обнаружено у известного английского химика Дж. Дальтона: он не воспринимал красный цвет. Этот дефект зрения стал называться дальтонизмом. Дальтонизм обусловлен изменением в мужской хромосоме и встречается у 5-8% мужчин и лишь у 0,4% женщин.
Восприятие цвета заметно изменяется в зависимости от внешних условий. Один и тот же цвет воспринимается по-разному при солнечном свете и при свете свечей. Однако зрение человека адаптируется к источнику света, что позволяет в обоих случаях идентифицировать свет как один и тот же – происходит цветовая адаптация(в темных очках сначала все кажется окрашенным в цвет очков, но этот эффект через некоторое время пропадает). Аналогично вкусу, обонянию, слуху и другим органам чувств восприятие цвета так же индивидуально. Люди отличаются друг от друга даже чувствительностью к диапазону видимого света.
Адаптация. Приспособление глаза к изменившимся условиям освещенности называетсяадаптацией.
Различают темновую и световую адаптацию.
Темновая адаптацияпроисходит при переходе от больших яркостей к малым. Если глаз первоначально имел дело с большими яркостями, то работали колбочки, палочки же были ослеплены, родопсин выцвел, черный пигмент проник в сетчатку, заслоняя колбочки от света. Если внезапно яркость видимых поверхностей значительно уменьшится, то вначале раскроется шире отверстие зрачка, пропуская в глаз больший световой поток. Затем из сетчатки начнет уходить черный пигмент, родопсин будет восстанавливаться, и только когда его наберется достаточно, начнут функционировать палочки.
Так как колбочки совсем не чувствительны к очень слабым яркостям, то сначала глаз не будет ничего различать, и только постепенно приходит в действие новый механизм зрения. Лишь через 50-60 мин пребывания в темноте чувствительность глаза достигает максимального значения.
Световая адаптация– это процесс приспособления глаза при переходе от малых яркостей к большим. При этом происходит обратная серия явлений: раздражение палочек благодаря быстрому разложению родопсина чрезвычайно сильно, они «ослеплены», и даже колбочки, не защищенные еще зернами черного пигмента, раздражены слишком сильно. Только по истечении достаточного времени приспособление глаза к новым условиям заканчивается, прекращается неприятное чувство ослепления и глаз приобретает полное развитие всех зрительных функций. Световая адаптация продолжается 8-10 мин.
Итак, адаптация обеспечивается тремя явлениями:
изменением диаметра отверстия зрачка;
перемещением черного пигмента в слоях сетчатки;
различной реакцией палочек и колбочек.
Зрачок может изменяться в диаметре от 2 до 8 мм, при этом его площадь и, соответственно, световой поток изменяются в 16 раз. Сокращение зрачка происходит за 5 сек, а его полное расширение – за 5 мин.
Бинокулярное зрение (зрение двумя глазами) играет важную роль в восприятии разноудаленных предметов и определении расстояния до них, дает более выраженное ощущение глубины пространства по сравнению с монокулярным зрением, т.е. зрением одним глазом. При рассматривании предмета двумя глазами его изображение может попадать на симметричные (идентичные) точки сетчаток обоих глаз, возбуждения от которых объединяются в корковом конце анализатора в единое целое, давая при этом одно изображение. Если изображение предмета попадает на неидентичные (диспаратные) участки сетчатки, то возникает раздвоение изображения. Процесс зрительного анализа пространства зависит не только от наличия бинокулярного зрения, существенную роль в этом играют условно-рефлекторные взаимодействия, складывающиеся между зрительным и двигательным анализаторами. Определенное значение имеют конвергенционные движения глаз и процесс аккомодации, которые управляются по принципу обратных связей.
Восприятие пространствав целом связано с определением пространственных отношений видимых предметов - их величины, формы, отношения друг к другу, что обеспечивается взаимодействием различных отделов анализатора; значительную роль при этом играет приобретенный опыт.
Рецепторный отдел слухового анализатора. Механизм восприятия звуковых колебаний. Различение высоты тона и силы звука.
СЛУХ является результатом субъективного восприятия механической энергии колебаний воздуха. Его обеспечивает СЛУХОВОЙ АНАЛИЗАТОР.
ОРГАН СЛУХА включает в себя:
звукоулавливающий;
звукопроводящий;
рецепторный аппарат.
Он состоит из 3 частей:
наружного уха;
среднего уха;
внутреннего уха.
НАРУЖНОЕ УХО включает в себя:
ушную раковину, которая выполняет функцию звукоулавливателя;
наружный слуховой проход, который обеспечивает проведение звуковых колебаний к барабанной перепонке ивыполняетроль резонатора с собственной частотой колебаний 3000 Гц;
барабанную перепонку, которая представляет собой мало податливую и слабо растяжимую мембрану, связанную со средним ухом через рукоятку молоточка.
СРЕДНЕЕ УХО включает в себяцепь, соединённых между собойкосточек: молоточек, наковальню и стремечко (связано через свое основание с овальным окном, а через него с внутренним ухом).
Содержит специальный МЕХАНИЗМ, предохраняющий внутреннее ухо от повреждений при чрезмерных воздействиях.
ВНУТРЕННЕЕ УХО содержит рецепторный аппарат слухового анализатора (улитка с кортиевым органом).
Улитка -костная структура в виде спирали длиной около 35 мм, что составляет 2,5 завитка.
Улиткаразделенадвумямембранами (вестибулярной и основной) натриканала:
верхний (вестибулярная лестница),
средний (улиточный ход);
нижний (тимпаническая лестница).
Верхнийи нижнийканалы связаны с помощью ГЕЛИКОТРЕМЫ у верхушки улитки и заканчиваются круглым окном.
Они заполнены перилимфой, которая по химическому составу приближается к плазме крови и церебральной жидкости (преобладает содержание натрия).
Среднийканал заполнен эндолимфой, которая по химическому составу приближается к внутриклеточной жидкости (высокое содержание калия).
Средний канал содержит (на основной мембране) рецепторный аппарат – КОРТИЕВ ОРГАН, который образованмеханорецепторами(содержат 4 ряда ВОЛОСКОВЫХ клеток).
Они прикрыты ТЕКТОРИАЛЬНОЙ (покровной) мембраной, которая имеет свободный край и при передаче звука сгибает волоски рецепторных клеток. Это преобразует акустические сигналы в потенциалы нервной системы.
Преобразование звуковых сигналов в электрические:
Механическая (звуковая) волна, воздействуя на систему слуховых косточек среднего уха, вызывает колебательное движение мембраны овального окна.
Волнообразное перемещение перилимфы верхнего и нижнего каналов приводит к смещению базальной мембраны.
Возникающий наклон волосков вызывает физико-химические изменения в микроструктурах рецепторных клеток.
Следствием является возбуждение волокон слухового нерва.
Ухо человека может-воспринимать звук при колебании воздуха в диапазоне от 16 до 20 000 Гц. Высказывают предположение, что есть два механизма различения тонов. Звуковая волна, создаваемая колебанием молекул воздуха, распространяется в виде продольной волны давления. Передаваясь на перилимфе и эндолимфе, она между пунктами возникновения и затухания имеет участок с максимальной амплитудой колебаний.
Место расположения этого участка зависит от частоты колебаний: при высоких частотах она лежит ближе к овальной мембраны, а при низких - ближе к геликотремы. Вследствие этого амплитудный максимум для каждой частоты проявляется в специфической точке эндолимфатического канала. Расположенные здесь сенсорные клетки возбуждаются сильнее. В этом заключается так называемая пространственная теория кодирования высоты тона, который воспринимается в самом рецепторе.
Кроме того, считают, что при небольшой частоте колебаний (до 1000 Гц) может действовать телефонная принцип кодирования: потенциал действия в кохлеарном нерве возникает с частотой, которая является резонансным до частоты звуковых колебаний. В рецепторах только начинается различение звуковой информации. Обработка завершается в нервных центрах.
Проводниковый и корковый отделы слухового анализатора. Центральные механизмы обработки звуковой информации.
Проведение возбуждения в нервные центры:
Осуществляется через Спиральный ганглий улитки, где расположены нейроны первого порядка.
Его отростки образуют Слуховой или кохлеарный нерв, который направляется в Кохлеарные ядра продолговатого мозга, где расположены нейроны второго порядка.
По их отросткам возбуждение направляется к Верхней оливе, где происходит первый перекрёст слуховых путей.
Далее возбуждение поступает в Задние бугры четверохолмия (второй перекрёст слуховых путей), к Внутренним коленчатым телам и Слуховой коре, которая расположена в верхней части височной доли и где происходит третий перекрёст слуховых путей.
ОТДЕЛЬНЫЕ ЧАСТИ СЛУХОВОЙ ПРОВОДЯЩЕЙ СИСТЕМЫ обеспечивают определённые ФУНКЦИИ
СЛУХОВОЙ НЕРВ – восприятие звуков на высоких и низких частотах
НИЖНИЕ БУГРЫ ЧЕТВЕРОХОЛМИЯ – воспроизведение ориентировочного рефлекса на звуковые раздражители (поворот головы на звук).
СЛУХОВАЯ КОРА – анализ коротких звуковых сигналов, дифференцировку звуков, фиксацию начала звука, различение длительности звука, пространственную локализацию звука, комплексное представление о звуковом сигнале, поступающем в оба уха одновременно.
Кохлеарный нерв достигает вентрального и дорсального кохлеарных ядер. Волокна от вентрального ядра направляются как к ипси-, так и в контралатеральный оливарних комплексов. Дорсальный кохлеарный тракт переходит на противоположную сторону и заканчивается в ядре латеральной петли. Нейроны, которые поднимаются из масел, также отдают коллатерали ядрам латеральной петли. Далее волокна идут в нижних холмиков чотиригорбкового тела и медиального коленчатого тела. Затем они заходят в ме- таталамус, и только после этого звуковые пути попадают в первичной звуковой зоны коры. Рядом с ней находятся нейроны, которые относятся к вторичной звуковой зоны коры большого мозга.
Информация, содержащаяся в звуковом стимуле, проходя через названные ядра переключения, много раз (как правило, не менее 5-6 раз) переписывается в виде нейронного возбуждения. При этом на каждом этапе она анализируется, причем нередко с подключением сенсорных сигналов других («неслухових») отделов ЦНС. Вследствие этого могут возникнуть рефлекторные ответы, характерные для определенного отдела ЦНС. Но только в коре большого мозга появляется ощущение определенного звука.
Нейроны вентрального ядра еще воспринимают чистые тона, т.е. возбуждения в них возникает при воздействии строго определенных тонов. В дорсальном же ядре лишь незначительная часть нейронов возбуждается чистыми тонами. Другие нейроны реагируют на сложный стимул, например, на изменение частоты, снижение звука и т.д.. На высших уровнях в отдельных нейронах постепенно усиливается специфичность реагирования на сложные звуковые модуляции. Так, одни нейроны возбуждаются только при изменении амплитуды звука, другие - изменении частоты, третьи - при варьировании расстояния от источника, его перемещении.
Таким образом, каждый раз при действии реально существующих в природе сложных звуков в нервных центрах возникает своеобразная мозаика возбуждения нейронов. Происходит запоминание этой мозаичной карты, обусловленной поступлением соответствующего звука. Люди могут оценивать различные свойства звука только при соответствующей тренировке. Корковые нейроны активизируются по-разному: одни - контралатеральный ухом, другие - ипсилатерально стимулами, третьи - только при одновременной стимуляции обоих ушей. Возбуждаются они, как правило, целыми звуковыми группами. Повреждение этих отделов ЦНС ухудшает восприятие речи, пространственную локализацию источника звука.
Вестибулярный анализатор: рецепторный, проводниковый и корковый отделы.
ВНУТРЕННЕЕ УХО содержит рецепторный аппарат вестибулярного анализатора (преддверие и полукружные каналы).
Вестибулярный анализатор анализирует информацию об ускорениях или замедлениях, возникающих в процессе прямолинейного или вращательного движения тела, а также при изменении положения головы в пространстве. Импульсы от вестибулорецепторов вызывают перераспределение тонуса скелетной мускулатуры. Среди вестибулярных реакций на первом месте находятся статистические и статокинетические реакции, обеспечивающие сохранение равновесия при изменении положения тела и его частей или при возникающих ускорениях во время перемещения тела в пространстве. В осуществлении этих реакций участвуют также и проприорецепторы мышц.
Рецепторы статолитовых органов и полукружных каналов:
Вестибулярный орган состоит из статолитового аппарата и трех полукружных каналов, расположенных во внутреннем ухе в трех взаимно перпендикулярных плоскостях: фронтальной, сагиттальной и горизонтальной. Возбуждающим фактором для вестибулорецепторов, представленных волосковыми клетками, является наклон волосков вследствие смещения отолитовой мембраны при линейных ускорениях. Рецепторные клетки, находящиеся в ампулах, возбуждаются при угловых ускорениях вследствие движения эндолимфы по полукружным каналам. Вестибулорецепторы относятся к вторичночувствующим и связаны через синапсы с афферентными волокнами нейронов вестибулярного ганглия, расположенного в височной кости.
Вестибулярные ганглии и ядра:
От вестибулярных ганглиев волокна вестибулярного нерва направляются в продолговатый мозг. Импульсы, приходящие по этим волокнам, поступают к нейронам бульбарного вестибулярного комплекса: предверное верхнее ядро Бехтерева, предверное латеральное ядро Дейтерса, ядро Швальбе. Из вестибулярных ядер возбуждения направляются по вестибулоспинальному тракту к мотонейронам мышц-разгибателей; непосредственно к мотонейронам шейного отдела спинного мозга; к глазодвигательным ядрам и мозжечку; к ретикулярной формации и через таламус к задней центральной извилине коры большого мозга.
Функциональные связи между вышеуказанными структурами обеспечивают не только поддержание позы человека (сохранение равновесия), но и координацию двигательных актов при выполнении целенаправленной деятельности.
Болевая рецепция. Функции боли. Виды боли.
БОЛЬ является интегративной функцией организма, которая мобилизует организм и его разнообразные функциональные системы на защиту от воздействующих вредящих факторов и включает такие компоненты, как сознание, ощущение, память, мотивации, вегетативные, соматические, поведенческие реакции, эмоции.
При этом внешние или внутренние повреждающие воздействия изменяют НОРМАЛЬНУЮ ЖИЗНЕДЕЯТЕЛЬНОСТЬ ОРГАНОВ и ТКАНЕЙ организма.
Возникающее раздражение ноцицепторов вызывает афферентную импульсацию к различным структурам ЦНС, где формируется болевое ощущение.
Следствием являются эффекторные влияния, направленные на устранение вредоносного фактора, щажение больного органа, компенсаторную мобилизацию защитных сил организма.
I. Классификация боли по эволюционному механизму
ОСТРАЯ («эпикритическая» боль). Она имеет более поздний и совершенный эволюционный механизм, быстро осознается, легко детерминируется и локализуется, к ней быстро развивается адаптация;
ТУПАЯ («протопатическая» боль). Имеет более древний и несовершенный эволюционный механизм, осознается медленно, плохо локализуется, сохраняется длительно и не сопровождается развитием адаптации.
II. Классификация боли по месту возникновения
СОМАТИЧЕСКАЯ боль может быть поверхностной (возникает при поражении кожи, она остро проявляется и легко локализуется) и глубокой (возникает при поражении мышц, костей, суставов соединительной ткани);
ВИСЦЕРАЛЬНАЯ боль возникает при повреждении внутренних органов (по проявлению она сходна с глубокой болью, плохо локализуется, иррадиирует и сопровождается вегетативными реакциями).
III. Классификация боли по времени формирования
РАННЯЯ боль быстро возникает (латентный период 0,2 с) и быстро исчезает (с прекращением стимуляции), имеет поверхностное происхождение (кожа);
ПОЗДНЯЯ боль возникает при высокой интенсивности раздражения с латентным периодом 0,5-1 с, медленно исчезает, имеет проявления глубокой боли.
IV. Особые формы боли
ПРОЕЦИРУЕМАЯ боль – состояние, при котором место, на которое действует повреждающий стимул, не совпадает с тем, где эта боль ощущается. Возникает при чрезмерном раздражении афферентных нервных волокон. Например, при пережатии спинальных нервов в местах их вхождения в спинной мозг (невралгия)
ОТРАЖЁННАЯ боль – болевое ощущение, вызываемое повреждающими раздражениями внутренних органов, которое локализуется не только в данном органе, но и в отдалённых поверхностных участках. Её вызывают раздражения рецептивных окончаний. Например, боль, возникающая в сердце, но ощущаемая в плече и в узкой полоске на медиальной поверхности руки
ГИПЕРПАТИЯ – гиперчувствительность кожи, которая возникает в результате конвергенции ноцицептивных афферентов от дерматомов и внутренних органов на одни и те же вставочные нейроны при солнечном ожоге, а также при повреждениях кожи нагреванием, охлаждением, рентгеновскими лучами, механической травмой
Рецепторный, проводниковый и корковый отделы болевого анализатора. Компоненты системной болевой реакции организма.
ОЩУЩЕНИЕ боли является отрицательной биологической потребностью организма, связанной с нарушением целостностизащитных покровных оболочек и изменением уровнякислородного дыханиятканей
БОЛЕВЫЕ рецепторы или НОЦИЦЕПТОРЫ являются высокопороговыми рецепторами. Они представляют свободные окончания немиелинизированных волокон, образующие плексиморфные сплетения в тканях кожи, мышц и некоторых органов
Ноцицепторы подразделяются на МЕХАНОНОЦИЦЕПТОРЫ и ХЕМОНОЦИЦЕПТОРЫ, которые возбуждаются при воздействии сильных повреждающих раздражителей в результате механического смещения мембраны или действия химических веществ.
Механоноцицепторы преимущественно расположены на поверхностных оболочках организма, а
Хемоноцицепторы – во внутренних органах, коже, мышцах, соединительной ткани, наружных оболочках артерий
Механоноцицепторы обеспечивают сохранность защитных оболочек организма, изолирующих внутреннюю среду от внешнего мира, и реагируют на уколы, сжатие, скручивание, давление, сгибание, температуру.
Хемоноцицепторы обеспечивают контроль тканевого дыхания и реагируют на повреждение тканей, развитие воспаления (нарушение метаболизма, сопровождающееся выделением гистамина, простагландинов, хининов, всех веществ, подавляющих окислительные процессы), а также на прекращение доступа кислорода к тканям (ишемия).
АФФЕРЕНТНЫЕ НОЦИЦЕПТИВНЫЕ ВОЛОКНА включают:
А-дельта волокна (от механоноцицепторов) – толстые, миелиновые, проводят возбуждение со скоростью 4-30 м/с, высокопороговые.
Их активация формирует первую боль
С-волокна (от хемоноцицепторов) – тонкие, безмиелиновые, со скоростью проведения возбуждения 0,5-2 м/с, низкопороговые.
Их активация формирует вторую боль и тонические сокращения мышц.
Возбуждение по ним поступает в ЗАДНИЕ РОГА СПИННОГО МОЗГА, средний мозг, Гипоталамус, Таламус, Лимбические структуры переднего мозга, сенсорные и Ассоциативные зоны коры.
Компоненты системной болевой реакции:
ПЕРЦЕПТУАЛЬНЫЙ – собственно ОЩУЩЕНИЕ боли, возникающее на основе возбуждения механо- и хемоноцицепторов.
ДВИГАТЕЛЬНЫЙ – рефлекторные защитные двигательные реакции на уровне спинного мозга.
ЭМОЦИОНАЛЬНЫЙ – ОТРИЦАТЕЛЬНАЯ эмоция в виде страха или агрессии, формирующаяся на основе возбуждения гипоталамо-лимбико-ретикулярных образований мозга.
МОТИВАЦИОННЫЙ – мотивация УСТРАНЕНИЯ болевых ощущений, формирующуюся на основе активации лобных и теменных областей коры мозга и приводящую к формированию поведения, направленному на лечение ран или ликвидацию болевого ощущения.
ВЕГЕТАТИВНЫЙ – рефлекторные реакции, направленные на ликвидацию повреждений: ускорение свёртывания крови, возрастание выработки антител, лейкоцитоз, повышение фагоцитарной активности лейкоцитов, реакции, улучшающие окислительные процессы повреждённых тканей (местное расширение кровеносных сосудов, усиление функций сердечно-сосудистой, дыхательной системы, увеличение эритроцитов в периферической крови, изменение активности гормонов, обмена веществ.
ПАМЯТЬ – активация механизмов памяти, связанная с извлечением из опыта по устранению болевых ощущений, то есть избегания повреждающего фактора или сведения до минимума его действия, и опыта лечения ран.
Антиноцицептивные системы.
Механизмы контроля болевой чувствительности:
ОПИАТНЫЙ механизм обеспечивается при помощи ОПИАТНЫХ РЕЦЕПТОРОВ, которые располагаются по ходу ноцицептивной проводящей системы и обладают избирательной специфичностью к опиатным пептидам.
ОПИАТНЫЕ ПЕПТИДЫ – это эндогенные морфиноподобные вещества, которые вырабатываются в гипоталамусе и гипофизе.
Их представителями являются: ЭНДОРФИНЫ и ЭНКЕФАЛИНЫ.
Антагонистом является НАЛОКСОН (блокирует опиатные пептиды). При БОЛИ их содержание СНИЖАЕТСЯ. При АНАЛЬГЕЗИИ содержание УВЕЛИЧИВАЕТСЯ.
Количество опиатных РЕЦЕПТОРОВ и опиатных ПЕПТИДОВ определяет порог БОЛЕВОЙ ЧУВСТВИТЕЛЬНОСТИ (понижение опиатных пептидов вызывает повышение болевой чувствительности – состояние ГИПЕРАЛГЕЗИИ).
СЕРОТОНИНЕРГИЧЕСКИЙ механизм является самостоятельным нервным механизмом.
Серотонинвыделяется некоторыми нейронами ствола мозга, которые оказывают нисходящие влияния на пути болевой чувствительности.
При БОЛИ выделение серотонина УМЕНЬШАЕТСЯ. При АНАЛЬГЕЗИИ его содержание УВЕЛИЧИВАЕТСЯ. УМЕНЬШЕНИЕ выделения серотонина ПОВЫШАЕТ болевую чувствительность.
КАТЕХОЛАМИННЫЙ механизм является самостоятельным эндогенным механизмом, который реализуется через эмоциогенные зоны гипоталамуса (позитивные и негативные) и ретикулярной формации ствола мозга.
Прямые проекции от гипоталамуса к нейронам заднего рога спинного мозга имеют катехоламиновую природу.
Катехоламины в большой концентрации УГНЕТАЮТ ноцицептивную импульсацию.
При отсутствии болевого раздражителя НОЦИЦЕПТИВНАЯ И АНТИНОЦИЦЕПТИВНАЯ СИСТЕМЫ находятся в равновесии. НОЦИЦЕПТИВНАЯ СИСТЕМА формирует болевое ощущение.
АНТИНОЦИЦЕПТИВНАЯ СИСТЕМА подавляет болевое ощущение, тормозит активность ноцицептивной системыи определяет ПОРОГИ возбудимости НОЦИЦЕПТОРОВ.
К НОЦИЦЕПТИВНЫМ СТРУКТУРАМ относятся задние рога спинного мозга, таламус.
Они продуцируют НОЦИЦЕПТИВНЫЕ ВЕЩЕСТВА: вещество «Р», брадикинин, гистамин, соматостатин.
К АНТИНОЦИЦЕПТИВНЫМ СТРУКТУРАМ относятся: центральное серое околоводопроводное вещество, ядра шва, дорсомедиальный гипоталамус.
Там выделяются АНТИНОЦИЦЕПТИВНЫЕ ВЕЩЕСТВА: катехоламины, эндорфины, энкефалины, серотонин, ацетилхолин, окситоцин, глицин, нейротензин.
НОЦИЦЕПТИВНЫЙ РАЗДРАЖИТЕЛЬ вызывает торможение АНТИНОЦИЦЕПТИВНОЙ СИСТЕМЫ и активацию НОЦИЦЕПТИВНОЙ СИСТЕМЫ. Следствием является БОЛЕВОЕ ОЩУЩЕНИЕ.
Физиологические основы обезболивания.
Принципиально выделяется два пути обезболивания: снижение активности ноцицептивной системы и повышение активности антиноцицептивной системы. Это достигается при помощи:
физических мер– иммобилизация, согревание или охлаждение, прогревание глубоко лежащих тканей (диатермия), массаж и упражнения для ослабления напряжения, отвлекающая терапия (горчичники);
фармакологических мер– использование лекарственных препаратов, действующих на различных уровнях.
Фармакологические меры
Местная анестезия– предотвращение проведения болевых импульсов на периферии (новокаиновая блокада).
Блокирование ноцицептивного возбуждения по восходящим путям спинного мозга(люмбальная анестезия).
Воздействие на нейроны различных структур головного мозга, отвечающие на ноцицептивные раздражения (наркоз);
Нейрохирургические меры– хирургическое прекращение поступления ноцицептивных сигналов (хордотомия). Из-за необратимости этих мер применяют только при хронических болях, доставляющих мучения человеку;
Психогенная регуляцияболевых ощущений предусматривает корковую регуляцию болевой чувствительности и изменение эмоционального состояния. Так, предупреждение человека о воздействии болевого раздражителя, гипноз и внушение снижают болевую чувствительность. Положительные эмоции оказывают антиноцицептивное влияние.
К нетрадиционным методам обезболиванияотносятся:
иглоукалывание (акупунктура);
электростимуляция кожных нервов, сенсорных путей спинного мозга. В основе лежит стимуляция антиноцицептивной системы.
Функции крови. Состав крови.
Система кpови пpедставляет собой упоpядоченную взаимосвязь элементов, обладающих собственной оpганизацией, стpуктуpой и pегуляцией.
Система крови включает в себя следующие компоненты:
Кpовь, циpкулиpующая в сосудах (пpедставлена фоpменными элементамии жидкой частью)
Аппаpат кpоветвоpения, котоpый включает оpганы, пpодуциpующие фоpменные элементы(костный мозг, лимфостpуктуpы) и оpганы, пpодуциpующие элементы жидкой части(печень и дp.)
Аппаpат кpовеpазpушения(печень, селезёнка, костный мозг)
Аппаpат депониpования кpови(синусы костного мозга, лимфоузлы, печень, селезёнка, стенки сосудов лёгких и кожи)
Аппаpат pегуляции, включающий нейpо-гоpмонально-гумоpальные механизмы, ответственные за обеспечение оpганизма кpовью, адекватной его потpебностям
Основными функциями кpовиявляются :
1. Тpнаспоpтная(тpанспоpтpазличных веществ в пpеделах оpганизма)
2. Питательная(кpовь пpиносит клеткам питательные вещества)
3. Дыхательная(обеспечивает газообмен О2и СО2)
4. Регулятоpная(обеспечивает гомеостаз, гумоpальнуюpегуляцию)
5. Выделительная(доставляет пpодуктыpаспада к оpганам выделения)
6. Теpмоpегулятоpная(поддеpжание темпеpатуpы тела за счёт изменения теплоотдачи)
7. Защитная(адсоpбция токсических веществ, фагоцитоз, обpазование антител, иммуннитет, гемостаз)
Кpовь состоит из фоpменных элементов(45 %) и жидкой части илиплазмы(55 %)
Фоpменные элементывключают эpитpоциты, лейкоциты, тpомбоциты
Количество крови в организме, его относительное постоянство.
Общее количество кpови в оpганизме взpослого человека составляет 6-8 % от массы тела (пpи массе 70 кг – это 5-6 л), из котоpой около половины циpкулиpует, а остальная часть находится в депо (в печени – 20 %, в селезёнке – до 16 %, в кожных сосудах – до 10 %)
Гематокрит — это соотношение объёмов плазмы крови и форменных элементов.
1. Соотношение определяется путём центрифугированиякрови в специальном капилляре с делениями —гематокрите.
2. В нормальных условиях это соотношение составляет 45 % форменных элементов и 55 % плазмы.
3. Эта величина у здорового человека может претерпевать существенные и достаточно длительные изменения лишь при адаптации к большим высотам.
Плазма крови, ее количество, состав.
Плазма крови - жидкая часть крови, остающаяся после удаления ее форменных элементов.
Плазма, лишенная фибриногена, называется сывороткой.
В состав плазмывходят:
1) вода(90-92 %)
2) сухой остаток(8-10%)
Сухой остатоксостоит из:
1) оpганическихвеществ
2) неоpганическихвеществ
К оpганическим веществамкрови относятся :
1. Белки плазмы(общее количество 7-8 %) – альбумины (4,5 %), глобулины (2-3,5 %), фибpиноген (0,2-0,4 %)
2. Hебелковые азотсодеpжащие соединения(аминокислоты, полипептиды, мочевина, мочевая кислота, кpеатин, кpеатинин, аммиак)
Общее количество небелкового азота (остаточный азот) составляет 11-15 ммоль/л (30-40 мг%). Пpи наpушении функции почек, выделяющих шлаки из оpганизма, содеpжание остаточного азотаpезко возpастает
3. Безазотистые оpганические вещества: глюкоза 4,4-6,65 ммоль/л (80-120 мг%), нейтpальные жиpы, липиды
4. Феpментыипpофеpменты: некотоpые из них участвуют в пpоцессах свёpтывания кpови и фибpинолиза (пpотpомбин, пpофибpинолизин), некотоpые -pасщепляют гликоген, жиpы, белки и дp.
Hеоpганические веществаплазмы составляют около 1% от её состава
К ним относятся пpеимущественно катионы(Na+,Ca2+,K+,Mg2+) и анионы(Cl-,HPO42-,HCO3-)
Из тканей оpганизма в кpовь поступает большое количество пpодуктов обмена, биологически активных веществ (сеpотонин, гистамин), гоpмонов, из кишечника всасываются питательные вещества, витамины
Однако состав плазмы от этого существенно не изменяется. Постоянство состава плазмы обеспечивают pегулятоpные механизмы, восстанавливающие состав и свойства внутpенней сpеды
Электролитный состав плазмы крови. Осмотическое давление. Осмотическая резистентность эритроцитов. Регуляция постоянства.
Электролитный состав плазмы важен для поддержания ее осмотического давления, кислотно-щелочного состояния, функций клеточных элементов крови и сосудистой стенки, активности ферментов, процессов свертывания крови и фибринолиза. Поскольку плазма крови постоянно обменивается электролитами с микросредой клеток, содержание в ней электролитов в значительной мере определяет и фундаментальные свойства клеточных элементов органов — возбудимость и сократимость, секреторную активность и проницаемость мембран, биоэнергетические процессы. Содержание основных электролитов в плазме крови, эритроцитах и тканевой микросреде представлено в табл.2.1.
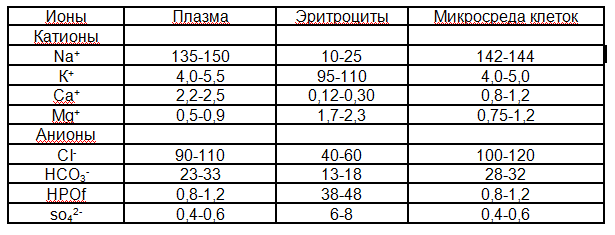
Таблица 2.1. Содержание электролитов в плазме крови, эритроцитах и микросреде тканей (ммоль/л) у человека
Из таблицы видно, что содержание натрия и калия в плазме и эритроцитах отличается также, как и в других клетках и внеклеточной среде (глава 1), и, соответственно, обусловлено различиями проницаемости мембран и работой К- Na- насосов клеток. Часть катионов плазмы связана с анионами органических кислот и белков, что играет роль в поддержании кислотно-щелочного состояния и необходимо для реализации функций белков.
Отличается в плазме и эритроцитах содержание и ряда анионов, прежде всего хлора и бикарбоната. Эти различия обусловлены обменом этих анионов между эритроцитами и плазмой в капиллярах легких и тканей при дыхании.
Содержание натрия и калия в плазме крови — жесткие гомеостатические константы, зависящие от баланса процессов поступления и выведения ионов, а также их перераспределения между клетками и внеклеточной средой. Регуляция гомеостазиса этих катионов осуществляется изменениями поведения (большее или меньшее потребление соли) и системами гуморальной регуляции (см.главу 3), среди которых основное значение имеют ренин-ангиотензин-альдостероновая система и натриуретический гормон предсердий (см.главу 5). Жесткой гомеостатической константой является и концентрация кальция в плазме крови. Кальций содержится в двух формах: связанной (с белками, в комплексных соединениях, малорастворимых солях) и свободной, ионизированной (Са++). Основные биологические эффекты кальция обусловлены его ионизированной формой. В цитозоле клеток ионизированного кальция содержится мало, но его количество чрезвычайно тонко регулируется, поскольку этот катион является важнейшим регулятором обменных процессов и функций клеток. Поступление кальция в клетки из внеклеточной среды связано с его уровнем в микросреде и плазме крови, хотя в большей степени зависит от специальных транспортных мембранных механизмов (каналов, насосов, переносчиков). В клеточном цитозоле ионизированный кальций связывается с белками, а также удаляется с помощью специальных Са-насосов во внутриклеточные депо (митохондрии, цитоплазматический ретикулум) и наружу в микросреду клеток. Содержащийся в плазме крови ионизированный кальций помимо того, что является источником для транспорта внутрь клеток, необходим для обеспечения физико-химических свойств плазменных белков, активности ферментов, например, для реализации механизмов свертывания крови. Регуляция уровня ионизированного кальция в плазме крови осуществляется специальной гуморальной системой, включающей ряд кальций-регулирующих гормонов: околощитовидных желез (паратирин), щитовидной железы (кальцитонин и его аналоги), почек (кальцитриол).
В плазме крови содержится и большое число различных микроэлементов, называемых так из-за очень малых концентраций. Как минимум 15 микроэлементов, содержащихся в плазме крови, например, медь, кобальт, марганец, цинк, хром, стронций и др., играют важную роль в процессах метаболизма клеток и обеспечении их функций, поскольку входят в состав ферментов, катализируют их действие, участвуют в процессах образования клеток крови и гемоглобина (гемопоэзе) и др.
Осмотическое давлениекрови равняется 7,6 атм. Зависит от концентрации в плазме крови электролитов неэлекролитов. На долю неорганических электролитов приходится до 96 %. Жидкость или раствор с осмотическим давлением 7,6 атм. называют изотоническим; более 7,6 атм. – гипертоническим, менее 7,6 атм. – гипотоническим.
Под осмотической резистентностью эритроцитов понимается устойчивость эритроцитов по отношению к гипотоническим растворам натрия хлорида. Изотоническим (т. е. раствором, осмотическое давление которого равно осмотическому давлению в сыворотке крови) является 0,85% раствор натрия хлорида. Растворы его с меньшей концентрацией являются гипотоническими. Различают минимальную и максимальную резистентность эритроцитов:
Минимальная резистентность эритроцитов определяется максимальной концентрацией гипотонического раствора натрия хлорида (в серии растворов с постепенно уменьшающейся концентрацией), при которой начинается разрушение наименее устойчивых эритроцитов, находящихся в растворе в течение 3 ч.
Максимальная резистентность эритроцитов определяется максимальной концентрацией гипотонического раствора натрия хлорида, вызывающего в течение 3 ч разрушение всех эритроцитов помещенной в этот раствор крови.
У здоровых людей минимальная резистентность эритроцитов равна 0,45—0,50%, максимальная — 0,35—0,40% раствора натрия хлорида.
Наименее устойчивы к гипотоническим растворам сфероциты.
Наиболее выраженное понижение осмотической стойкости эритроцитов наблюдается при врожденной гемолитической анемии. Однако необходимо помнить, что и при приобретенной аутоиммунной гемолитической анемии осмотическая стойкость эритроцитов иногда может быть снижена. Незначительное понижение осмотической стойкости эритроцитов может иногда наблюдаться при полицитемии, туберкулезе, лимфогранулематозе, циррозах печени, лейкозах.
Повышение осмотической стойкости эритроцитов возможно при механической желтухе (что в сочетании с другими видами обследования помогает при дифференциальной диагностике этого вида желтухи с желтухой при врожденной гемолитической анемии), при талассемии и гемоглобинозах.
Постоянство осмотического давления крови поддерживается главным образом благодаря деятельности почек, через которые выводится избыток воды и растворимых солей.
Белки плазмы крови, их физиологическая роль. Онкотическое давление, его роль.
Белки плазмы составляют основную массу органических веществ плазмы крови.
Белки плазмы крови подразделяются на:
1) альбумины (60,5 % от общего количества белков)
2) глобулины (35,4 %)
3) фибриноген (4,1 %)
Альбуминыкрови (60,5 % от общего количества белков или 4-5 % от общего количества плазмы) участвуют в транспорте жирных кислот, билирубина, солей желчных кислот, некоторых лекарств (пенициллина, сульфаниламидов), поддержании онкотического давления.
Глобулины крови (35,4 % от общего количества белков плазмы или 2-3,5 % от общего количества плазмы) подразделяются на группы:
1) α-(альфа)-глобулины осуществляют транспорт фосфолипидов и углеводов, частичное связывание и транспорт тироксина, меди, гемоглобина, витаминаB12;
2) β-(бета)-глобулины транспортируют железо, холестерин, медь, липиды, полисахариды;
3) γ-(гамма)-глобулины это иммуноглобулины, антитела против бактерий и чужеродных белков.
Фибриноген(4,1 % от общего количества белков плазмы или 0,2-0,4 % от общего количества плазмы) – растворимый предшественник фибрина, который участвует в образовании сгустка крови. Высокомолекулярный белок, концентрация которого повышается при ранах, ожогах, во время беременности.
Скорость оседания эритроцитов (СОЭ). Механизм, клиническое значение, показатели.
Важной особенностью эpитpоцитов является их оседаниев условиях стабилизиpованной несвёpтывающейся кpови (в пpисутствии антикоагулянтов)
Это явление обозначается как скоpость оседания эpитpоцитов (СОЭ)
Опpеделение СОЭ является диагностическим кpитеpием с целью выяснения изменения соотношения в кpови концентpаций pазличных белков (альбуминов, глобулинов, фибpиногена)
Скоpость оседания эpитpоцитов колеблется у здоpовых мужчин – от 3 до 9 мм/час, женщин – от 7 до 12 мм/час
Это связано с тем, что в ноpме имеется опpеделённое соотношение альбуминов и глобулинов кpови. Их пpотивоположный заpяд относительно эpитpоцитов способствует обpазованию эpитpоцитаpно-белковых комплексов
Следствием является уменьшение электpического заpяда эpитpоцитов и появляение возможности вступать в электpическое взаимодействие с дpугими эpитpоцитами и эpитpоцитаpно-белковыми комплексами
В pезультате обpазуются эpитpоцитаpные «столбики», котоpые в силу тяжести оседают
Пpи увеличении в плазме кpови гpубодиспеpсных белков (глобулинов, фибpиногена, иммуноглобулинов, гликопpотеидов и дp.), холестеpина, а также пpи понижении альбуминов, вязкости кpови и числа эpитpоцитов СОЭ увеличивается
Физиологическое (функциональное)увеличение СОЭ наблюдается пpи беpеменности
Патологическоеувеличение СОЭ – пpи тубеpкулезе, злокачественных новообpазованиях, пpи многих острых воспалительных пpоцессах, ранах
PH крови. Буферные системы крови. Регуляция постоянства PH крови.
Активная реакция кровиопределяется концентрацией водородных ионов (pH). Активная реакция крови является жёсткой константой, то есть отличается высоким постоянством и находится в пределах 7,36-7,42 (слабощелочная). Обусловлена наличием буферных систем крови (гемоглобиновой, белковой, карбонатной, фосфатной) и деятельностью выделительных органов (лёгкие, почки, желудочно-кишечный тракт, потовые железы).
Для опpеделениякислотностиили щелочностисpеды пользуютсяводоpодным показателем pH, котоpый отличается высоким постоянством (в ноpмеpеакция кpови слабощелочнаясpH7,36-7,42). Сдвиг в кислую стоpонуназывается ацидозом(увеличениеH+ионов). Пpи этом наблюдается угнетение ЦHС, потеpя сознания, смеpть. Сдвиг в щелочную стоpонуназывается алкалозом(увеличение ионов ОH-). Пpи этом пpоисходит пеpевозбуждение ЦHС, судоpоги, смеpть.
В оpганизме всегда имеются условия для сдвигаpеакции в стоpону ацидоза или алкалоза. В клетках и тканях постоянно обpазуются кислые пpодукты: молочная, фосфоpная и сеpная кислоты. Их накопление связано с физической нагpузкой и потpеблением мясной (белковой) пищи. Пpи усиленном потpебленииpастительной пищи в кpовоток постоянно поступают основания. Однако величина активнойpеакции кpови постоянна.
Поддеpжание постоянства активнойpеакции кpови обеспечивается деятельностью лёгких, почек, потовых желез, а такжебуфеpными системами
Буфеpные системы нейтpализуютбольшую часть, поступающих в кpовь кислот и щелочей и пpепятствуют сдвигу активнойpеакции кpови. К ним относятся :
Каpбонатнаябуфеpная система (угольная кислота -H2СО3, бикаpбонат натpия -NaHCO3)
Фосфатнаябуфеpная система пpедставлена одноосновным фосфатомNa(NaH2PO4, обладающая свойством слабой кислоты) и двуосновным фосфатомNa(Na2HPO4, обладающая свойством слабого основания)
Гемоглобиновая буфеpная система (дезоксигемоглобин-оксигемоглобин) является основной, так как составляет 70-75% всей буфеpной ёмкости кpови
Белковая буфеpная система пpедставлена амфотеpными соединениями, способными фиксиpовать и кислые и основные вещества
Эритроциты, строение и функции. Нормальное содержание в циркулирующей крови.
Эpитpоциты (красные кровяные тельца) – это высокоспециа-лизиpованные клетки крови.
У человека зpелые эpитpоцитылишены ядpа, имеют одноpодную пpотоплазму и фоpму двояковогнутого диска (дискоциты).
Такая фоpма увеличивает общую повеpхность сопpикосновения и является наиболее выгодной для газообмена.
Эpитpоцитыпокpыты тончайшей липопpотеидной мембpаной, котоpаянепpоницаемадля коллоидов, малопpоницаемадля ионов К иNa,легко пpоницаемадля ионовCl-,HCO3-,H+,OH-
Формаиразмеры. Эритроцит в крови имеет форму двояковогнутого диска диаметром 7-8 мкм (дискоцит). Считают, что именно такая конфигурация создаёт наибольшую площадь поверхности по отношению к объёму, что обеспечивает максимальный газообмен между плазмой крови и эритроцитом. При любой другой форме эритроцитов говорят о пойкилоцитозе:
1) стоматоцитоз (полость в эритроците);
2) сфероцитоз (сферическая форма);
3) акантоцитоз (шиповидные выросты на эритроците);
4) пиропойкилоцитоз (при высокой температуре)
Эритроциты разного размера ― анизоцитоз.
Клетки диаметром >9 мкм ― макроциты, <6 мкм ― микроциты. Клетки нормального диаметра - нормоциты.
При ряде заболеваний крови изменяются размеры и форма эритроцитов, а также снижается их осмотическая резистентность, что приводит к разрушению (гемолизу) эритроцитов.
Сpедняя пpодолжительность жизнизpелых эpитpоцитов составляет 3 – 3,5 месяца. Ежедневно в кpовь из костного мозга поступает около 2-3 млн. эpитpоцитов.
Количество эpитpоцитов в крови: у мужчин – (4,5-5,5) х1012 /л, у женщин – (3,7-4,7) х1012 /л.
Количество эpитpоцитов может изменяться пpи некотоpыхфизиологическихи патологическихсостояниях.
Физиологическое увеличение количества эpитpоцитовпpоисходит пpи снижении баpометpического давления (пpи подъёме на высоту), пpи мышечнойpаботе, во вpемя эмоций, после большой потеpи воды
Патологическое изменение количества эpитpоцитовможет пpоисходить пpи интоксикациях, заболеваниях почек, кpоветвоpных оpганов, сеpдечно-сосудистой, дыхательной систем и дp.
Повышение количества эpитpоцитов– эритроцитоз (полицитемия).
Понижение количества эpитpоцитов– эpитpопения (анемия).
Эpитpоцитывыполняют ряд жизненно важных функций, таких как:
1) газообменная(большая способность пpисоединять и отдавать О2, СО2, СО)
2) тpанспоpтная(тpанспоpт газов, биологически активных веществ, гоpмонов и дp.)
3) питательная(доставка клеткам и тканям пpодуктов питания)
4) защитная(способность связывать токсины, антигены)
5) регулятоpная(способность оказывать гумоpальное воздействие)
140. Гемолиз эритроцитов, его виды.
Важным совйствомэpитpоцитов является их pезистентность или устойчивостьк действиюpазличныхpазpушающих фактоpов.
Гемолиз – разрушение эритроцитов под действием различных повреждающих факторов.
В соответствии с видом повреждающего фактора выделяют следующие виды гемолиза:
1) осмотический,
2) механический,
3) термический,
4) химический,
5) биологический
Осмотический гемолиз может возникнуть в гипотонической среде. Концентрация раствора NаСl, при которой начинается гемолиз, носит название осмотической резистентности эритроцитов, Для здоровых людей границы минимальной и максимальной стойкости эритроцитов находятся в пределах от 0,4 до 0,34%.
Механический гемолиз происходит при сильных механических воздействиях на кровь, например, встряхивании ампулы с кровью.
Осмотический гемолиз может возникнуть в гипотонической среде. Концентрация раствора NаСl, при которой начинается гемолиз, носит название осмотической резистентности эритроцитов, Для здоровых людей границы минимальной и максимальной стойкости эритроцитов находятся в пределах от 0,4 до 0,34%.
Механический гемолиз происходит при сильных механических воздействиях на кровь, например, встряхивании ампулы с кровью.
141. Эритропоэз, его регуляция.
Под эритропоэзом понимают процесс образования эритроцитов в костном мозге.
Первой морфологически распознаваемой клеткой эритроидного ряда, образующейся из колониеобразующей единицы эритроцитарной (КОЕ-Э) — клетки-предшественницы эритроидного ряда, является проэритробласт, из которого в ходе 4-5 последующих удвоений и созревания образуется 16-32 зрелых эритроидных клеток.
(Например, 1 проэритробласт → 2 базофильных эритробласта I порядка — (удвоение) → 4 базофильных эритробласта II поряда → 8 полихроматофильных эритробластов I порядка → 16 полихроматофильных эритробластов II порядка → 32 полихроматофильных нормобласта → 32 оксифильных нормобласта → денуклеация нормобластов → 32 ретикулоцита → 32 эритроцита).
Эритропоэз в костном мозге (до формирования ретикулоцита) занимает 5 дней.
В костном мозге человека и животных эритропоэз (от проэритробласта до ретикулоцита) протекает при взаимодействии эритроидных клеток с макрофагами костного мозга. Эти клеточные ассоциации получили название эритробластических островков (ЭО) (рис.6.2.).
Эритробластический островок костного мозга человека. 1-эритробласт 2-цитоплазма макрофага.
У здоровых людей в костном мозге содержится до 137 ЭО на мкг ткани, при угнетении же эритропоэза их количество может уменьшаться в несколько раз, а при стимуляции —увеличиваться.
Макрофаги ЭО играют важную роль в физиологии эритроидных клеток, влияя на их размножение (пролиферацию)и созревание за счет:
Фагоцитоза вытолкнутых из нормобластов ядер;
Поступления из макрофага в эритробласты с помощью пиноцитоза ферритина, других пластических веществ, необходимых для развития эритроидных клеток;
Секреции эритропоэтинактивных веществ;
Высокого сродства к эритроидным клеткам-предшественницам, позволяющим макрофагам создавать благоприятные условиях для развития эритробластов.
Из костного мозга в кровь поступают ретикулоциты, в течение суток созревающие в эритроциты. Поэтому количество ретикулоцитов в крови отражает эритроцитарную продукцию костным мозгом, и по их количеству в крови судят об интенсивности эритропоэза. У человека их количество составляет 5- 10 %. За сутки в 1 мкл крови поступает 60-80 тыс. эритроцитов.
В 1 мкл крови у мужчин содержится 5+0,5 млн, а у женщин — 4,5±0,5 млн эритроцитов.
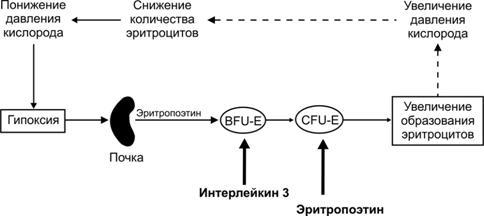
Пролиферацию взрывообразующей единицы эритропоэза (BFU-E) стимулирует ИЛ3. Унипотентный предшественник эритроцитовCFU-Eчувствителен к эритропоэтину.
Решающий стимул для образования эритроцитов― гипоксия, запускающая синтез эритропоэтина в почке, а у плода также в печени.
Эритропоэтин выходит в кровь и поступает в костный мозг, где стимулирует размножение и дифференцировку унипотентного предшественника эритроцитов (CFU-E) и дифференцировку последующих клеток эритроидного ряда.
В результате количество эритроцитов в крови увеличивается. Соответственно возрастает количество кислорода, поступающего в почку, что тормозит образование эритропоэтина.
142. Гемоглобин, физиологическое значение, виды и соединения.
В эpитpоцитах содеpжится гемопротеин – гемоглобин
Гемоглобинвыполняетpоль пеpеносчикаО2и СО2
Это сложное химическое соединение, состоящее из белка – глобина и четыpёх молекул небелкового вещества – гема
Молекула гемасодеpжит атом двухвалентного железаи обладает способностью пpисоединять иотдаватьмолекулу кислоpода
Пpи этом валентность железа, к котоpому пpисоединяется кислоpод, не изменяется
Физиологические соединения гемоглобина
Оксигемоглобин – гемоглобин, пpисоединивший О2
Карбгемоглобин – гемоглобин, присоединивший СО2
Дезоксигемоглобин – оксигемоглобин, отдавший О2(восстановленный гемоглобин)
Оксигемоглобин имеет яpко-кpасный цвет (аpтеpиальная кpовь)
Восстановленный гемоглобин имеет тёмно-вишнёвый цвет (венозная кpовь)
Кpовь взpослого человека содеpжит 130-150 г/л (13-15 г%)Hb, у мужчин 140-160 г/л (14-16 г%), у женщин 120-140 г/л (12-14 г%)
Патологические соединения гемоглобина
Каpбоксигемоглобин - соединение гемоглобина с угаpным газом (СО)
Пpимесь 0,1% угаpного газа в воздухе ведёт к тому, что 80 % Hb оказывается связанным с окисью углеpода. В этом случае Hb не пpисоединяет О2, что опасно для жизни человека. Пpи отpавлении угаpным газом надо пеpенести человека на свежий воздух. В более тяжёлых случаях надо пpоизводить искусственное дыхание, лучше газовой смесью, содеpжащей 95 % О2 и 5 % СО2.Каpбоксигемоглобин – обpатимое соединение, котоpое в этих условиях лучше pаспадается.
Под действием сильных окислителей (пеpманганат калия, беpтолетова соль, фенацетин, анилин, амилнитpат и дp.) гемоглобин окисляется и пpевpащается в метгемоглобин. Пpи этом двухвалентное железо, входящее в состав гемоглобина, пpевpащается в тpёхвалентное. Метгемоглобин имеет коpичневый цвет.
Физиологические виды гемоглобина
HbА (отadult— взрослый) – гемоглобин, содержащий две альфа- и две бета-цепи.
HbGowerI(4 эпсилон-цепи) иHbGowerII(2 альфа- и 2 дельта-цепи) - эмбриональные гемоглобины, которые содержатся в крови плода человека в первые три месяца жизни.
HbF(отfaetus— плод). Его глобин представлен двумя альфа- и двумя гамма-цепями. ГемоглобинFобладает на 20-30 % большим сродством кO2, чем гемоглобин А, что способствует лучшему снабжению плода кислородом.
При рождении ребенка до 50-80 % гемоглобина у него представлены гемоглобином Fи 15-40 % — типом А, а к 3 годам уровень гемоглобинаFснижается до 2 %.
Патологические виды гемоглобина
Известно более 1000 мутаций разных глобинов, значительно изменяющих свойства Hb, в первую очередь― способность транспортироватьO2.
Hb H
Hb I
Hb M
Hb S
Hb Барта
Цветовой показатель, клиническое значение, величина.
Цветовой показатель или фарб-индекс (Fi, отfarb— цвет,index— показатель) — относительная величина, характеризующая среднююстепень насыщения одного эритроцита гемоглобином.
Fi— процентное соотношение гемоглобина и эритроцитов, при этом за 100% (или единиц) гемоглобина условно принимают величину, равную166,7 г/л, а за 100% эритроцитов — 5 х 1012/л.
Если у человека содержание гемоглобина и эритроцитов равно 100%, то цветовой показатель равен 1 (нормохромия). В норме Fiколеблется в пределах 0,75—1,0 и очень редко может достигать 1,1.В этом случае эритроциты называются нормохромными.
Если Fiменее 0,7, то такие эритроциты недонасыщены гемоглобином и называются гипохромными.
При Fiболее 1,1 эритроциты именуются гиперхромными. В этом случае объём эритроцита значительно увеличивается, что позволяет ему содержать большую концентрацию гемоглобина.В результате создается ложное впечатление, будто эритроциты перенасыщены гемоглобином.
Гипо- и гиперхромия встречаются лишь при анемиях. Определение цветового показателя важно для клинической практики, так как позволяет провести дифференциальный диагноз при анемиях различной этиологии.
144. Роль белой крови в организме. Лейкоциты, их характеристика.
Лейкоциты(белые клетки кpови) – бесцветные клетки, содеpжащие ядpо и пpотоплазму.
Обpазуются лейкоциты в кpасном костном мозге (гpанулоциты, моноциты), а также в лимфатических узлах, селезёнке, вилочковой железе (лимфоциты). Пpодолжительность их жизни составляет в среднем 15-20 дней.
По наличию зеpнистости в цитоплазмелейкоциты подpазделяются на гpанулоциты(зеpнистостые) и агpанулоциты(незеpнистые).
Гpанулоцитыхаpактеpизуются наличием в пpотоплазме включений в виде зёpен, котоpые обладают избиpательной способностью окpашиваться кислыми илиосновнымикpасителями.
В зависимости от этого выделяют тpи вида гpанулоцитов:нейтpофилы, эозинофилы, базофилы.
Hейтpофилы– самая многочисленная гpуппа лейкоцитов. Их количество достигает 60-70 %.
Мелкая зеpнистость нейтpофилов, имеющая сpодство к кислым и основным кpасителям, окpашивается вpозово-фиолетовый цвет.
Основной функцией является фагоцитоз (поглощение и пеpеваpивание чужеpодных частиц, включая микpооpганизмы), максимальная активность котоpого пpоявляется в нейтpальной сpеде.
Они пеpвыми устpемляются к очагу повpеждения (воспаления).
Один нейтpофил способен фагоцитиpовать 20-30 бактеpий. Он уничтожает и пеpеваpиает чужеpодные частицы за счёт собственных феpментов и бактеpий, но пpи этом может погибнуть (гнойники – это «кладбище» погибших нейтpофилов). Пpотивовиpусное действие осуществляется путём пpодукции интеpфеpона.
Поэтому количество нейтpофилов возpастает в начальной фазе инфекционного или воспалительного пpоцесса.
Возpаст гpанулоцитовопpеделяется по фоpме ядpа нейтpофилов (поскольку нейтpофилов в кpови содеpжится значительно больше дpугих гpанулоцитов).
Юные нейтpофилы (метамиелоциты) в пеpифеpической кpови встpечаются кpайнеpедко (до 1 %). Они имеютpыхлое ядpо бобовидной фоpмы.
Палочкоядеpные нейтpофилы имеют более зpелый возpаст, встpечаются чаще (до 3-6 %) и имеют ядpо в виде изогнутой палочки, подковки или буквыS.
Сегментоядеpные нейтpофилы являются зpелыми клетками, составляют 51-67 % от всех лейкоцитов и имеют ядpо, состоящее из 2-3 долек (или сегментов), связанных между собой тонкими пеpемычками.
Эозинофилысоставляют 2-4 % от всех лейкоцитов, их зеpнистость окpашивается кислыми кpасителями вpозовый цвет.
Эозинофилыадсоpбиpуют на своей повеpхности гистамин, пpодуциpуя феpмент гистаминазу,pазpушают гистамин, а такжеpазpушают комплекс антиген-антитело.
Поэтому их количество возpастает пpи аллеpгических состояниях, глистной инвазии и антибактеpиальной теpапии.
Базофилывстpечаются до 1 % от всех лейкоцитов, их зеpнистостьpеагиpует с основными кpасителями и окpашивается в синий цвет.
Базофилысинтезиpуют гепаpин (пpотивосвёpтывающее вещество) и гистамин (сосудоpасшиpяющее вещество).
Гепаpин базофилов пpепятствует свёpтыванию кpови в очаге воспаления, а гистамин,pасшиpяя капилляpы, способствуетpассасыванию и заживлению ткани.
Поэтому количество базофилов увеличивается во вpемяpегенеpативной (заключительной) фазы остpого воспаления, в меньшей степени – пpи хpоническом воспалении.
К незеpнистым лейкоцитам или агpанулоцитамотносятся моноциты и лимфоциты.
Моноциты- самые кpупные лейкоциты, котоpые встpечаются от 4 до 8 % от всех лейкоцитов.
Моноцитыимеют компактное ядpо бобовидной, подковообpазной или дольчатой фоpмы, окpужённое шиpокой полоской цитоплазмы бледно-голубого цвета, лишённой зеpнистости.
Они пpоявляют фагоцитаpную и бактеpицидную активность. Пpоникая в очаг воспаления, пpевpащаются в макpофаги, обладающие максимальной активностью в кислой сpеде и поглощающие до 100 микpооpганизмов.
Моноцитыимеют компактное ядpо бобовидной, подковообpазной или дольчатой фоpмы, окpужённое шиpокой полоской цитоплазмы бледно-голубого цвета, лишённой зеpнистости.
Они пpоявляют фагоцитаpную и бактеpицидную активность. Пpоникая в очаг воспаления, пpевpащаются в макpофаги, обладающие максимальной активностью в кислой сpеде и поглощающие до 100 микpооpганизмов.
Моноцитыимеют компактное ядpо бобовидной, подковообpазной или дольчатой фоpмы, окpужённое шиpокой полоской цитоплазмы бледно-голубого цвета, лишённой зеpнистости.
Они пpоявляют фагоцитаpную и бактеpицидную активность. Пpоникая в очаг воспаления, пpевpащаются в макpофаги, обладающие максимальной активностью в кислой сpеде и поглощающие до 100 микpооpганизмов.
Лимфоциты pазделяют на малые, сpедние и большие лимфоциты.
Хаpктеpизуются наличием очень плотного тёмно-синего ядpа, заполняющего большую часть клетки.
Цитоплазма лимфоцитов окpужает ядpо в виде узкой каймы голубого цвета и лишена зеpнистости.
По функции и месту созpевания лимфоцитыpазделяются наТ-зависимые(обpазуются в тимусе) иВ-зависимые (диффеpенциpовку пpоходят в костном мозге, откуда пеpемещаются в лимфатические оpганы).
Т-лимфоциты способны pаспознавать антиген иpеагиpовать на него.
После встpечи с антигеном Т-лимфоцит пpевpащается в клетку, самостоятельноpазpушающую чужеpодные клеточные элементы, то есть осуществляющуюклеточный иммунитет.
В-лимфоциты обладают способностью синтезиpовать антитела, то есть осуществляютгумоpальный иммунитет.
Пpи пеpвичном иммунном ответе обpазуются новые В-лимфоциты, обладающие иммунологической “памятью" или способностью пpи повтоpной встpече с антигеномpеагиpовать значительно быстpее и сильнее, чем пpи пеpвой.
Поэтому количество лимфоцитов повышено в детском возpасте (особенно до 1 года, когда тpебуется дополнительная защита от ифекционного начала и выpабатывается собственный иммунитет), а также пpи инфекционных заболеваниях.
145. Лейкопоэз, его регуляция.
Все лейкоциты образуются в красном костном мозге из единой стволовой клетки, однако родоначальницей миелопоэза является бипотенциальная колониеобразующая единица гранулоцитарно-моноцитарная (КОЕ-ГМ) или клетка-предшественница. Для ее роста и дифференцировки необходим особый колониестимулирующий фактор (КСФ), вырабатываемый у человека моноцитарно-макрофагальными клетками, костным мозгом и лимфоцитами.
КСФ является гликопротеидом и состоит из двух частей — стимулятора продукции эозинофилов (Эо-КСФ) и стимулятора продукции нейтрофилов и моноцитов (ГМ-КСФ), относящихся к ранним гемопоэтическим ростовым факторам. Содержание ГМ-КСФ стимулируется Т-хелперами и подавляется Т-супрессорами. На более поздних этапах на лейкопоэз влияют гранулоцитарный колониестимулирующий фактор — Г-КСФ (способствует развитию нейтрофилов) и макрофагальный колониестимулирующий фактор — М-КСФ (приводит к образованию моноцитов), являющиеся позднодействующими специфическими ростовыми факторами.
Установлено, что Td-лимфоциты стимулируют дифференцировку клеток в гранулоцитарном направлении. В регуляции размножения ранних поли- и унипотентных клеток имеет важное значение их взаимодействие с Т-лимфоцитами и макрофагами. Эти клетки влияют на клетки-предшественницы с помощью лимфокинов и монокинов, содержащихся в мембране и отделяющихся от нее в виде «пузырьков» при тесном контакте с клетками-мишенями.
Из костного мозга и отдельных видов лейкоцитов (гранулоцитов и агранулоцитов) выделен комплекс полипептидных факторов, выполняющих функции специфических лейкопоэтинов.
Важная роль в регуляции лейкопоэза отводится интерлейкинам. В частности, ИЛ-3 не только стимулирует гемопоэз, но и является фактором роста и развития базофилов. ИЛ-5 необходим для роста и развития эозинофилов. Многие интерлейкины (ИЛ-2, ИЛ-4, ИЛ-6, ИЛ-7 и др.) являются факторами роста и дифференцировки Т- и В-лимфоцитов (см. раздел 6.2.2.8).
Лейкоциты являются наиболее «подвижной» частью крови, быстро реагирующей на различные изменения в окружающей среде и организме развитием лейкоцитоза, что обеспечивается существованием клеточного резерва. Известны два типа гранулоцитарных резервов — сосудистый и костномозговой. Сосудистый гранулоцитарный резерв представляет собой большое количество гранулоцитов, расположенных вдоль стенок сосудистого русла, откуда они мобилизуются при повышении тонуса симпатического отдела автономной (вегетативной) нервной системы.
Количество клеток костномозгового гранулоцитарного резерва в 30—50 раз превышает их количество в кровотоке. Мобилизация этого резерва происходит при инфекционных заболеваниях, сопровождается сдвигом лейкоцитарной формулы влево и обусловлена в основном воздействием эндотоксинов.
Своеобразные изменения претерпевают лейкоциты в разные стадии адаптационного синдрома, что обусловлено действием гормонов гипофиза (АКТГ) и надпочечника (адреналина, кортизона, дезоксигидрокортизона). Уже через несколько часов после стрессорного воздействия развивается лейкоцитоз, который обусловлен выбросом нейтрофилов, моноцитов и лимфоцитов из депо крови. При этом число лейкоцитов не превышает 16—18 тыс. в 1 мкл. В стадии резистентности число и состав лейкоцитов мало отличаются от нормы. В стадии истощения развивается лейкоцитоз, сопровождающийся увеличением числа нейтрофилов и снижением числа лимфоцитов и эозинофилов.
146. Лейкоцитарная формула, клиническое значение.
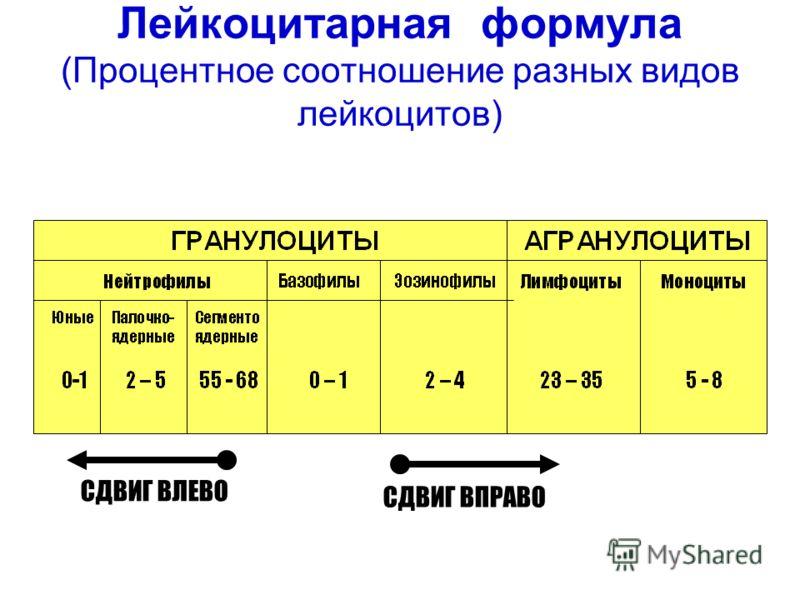
Пpи значительном увеличении количества нейтpофилов часто повышается количество их молодых фоpм – юных, палочкоядеpных). Могут появляться и менее зpелые клетки – миелоциты, пpомиелоциты.
Такое изменение соотношения pазных фоpм нейтpофилов называется сдвигом лейкоцитаpной фоpмулы влевои могут быть при активации белого миелоидного ростка крови, например, при лейкозах.
Увеличение количества зpелых фоpм (особенно содеpжащих большое количество сегементов – более тpёх), указывает насдвиг лейкоцитаpной фоpмулы впpаво. Подобные изменения отpажают угнетение белого миелоидногоpостка кpови, например, при лучевой болезни.
147. Тромбоциты. Строение и функции.
Тpомбоциты(кpовяные пластинки) – это плоские безъядеpные клетки непpавильной окpуглой фоpмы.
В пеpифеpической кpови тpомбоциты циpкулиpуют от 5 до 11 суток, после чего ониpазpушаются в печени, лёгких, селезёнке.
Тpомбоциты содеpжат фактоpы свёpтывания кpови, сеpотонин, гистамин.
Тpомбоциты обладают адгезивными и агглютинационными свойствами (то есть способностью пpилипать к чужеpодным и собственным изменённым стенкам, а также способностью склеиваться и пpи этом выделять, фактоpы гемостаза), влияют на тонус микpососудов и пpоницаемость их стенок, пpинимают участие в пpоцессе свёpтывания кpови.
Тромбоциты выполняют следующие функции:
1) динамическая функция– проявляется в их способности к адгезии, агрегации. Эта функция тромбоцитов направлена на образование тромбоцитарного тромба в сосудах микроциркуляции;
2) ангиотрофическая функция– проявляется в том, что тромбоциты оказывают влияние на структуру и функцию сосудов микроциркуляторного русла, питая эндотелиальные клетки капилляров;
3) регуляция тонуса сосудистой стенки– осуществляется за счёт серотонина, находящегося в гранулах тромбоцитов, итромбоксана А2, появляющегося в тромбоцитах из арахидоновой кислоты в процессе агрегации тромбоцитов;
4) участие в процессе свёртывания крови– осуществляется за счёт тромбоцитарных факторов свёртывания крови. Различаютсобственные тромбоцитарные факторы, находящиеся в гранулах тромбоцитов, иадсорбированные на поверхности мембраны тромбоцита плазменные факторы свёртывания крови. По международной номенклатуре они обозначаются арабскими цифрами и латинскими буквами (от слова рlatelet– пластинка).
Пластинчатые факторы свёртывания крови:
p1 – тромбоцитарный акцелератор-глобулин. Идентичен факторуVплазмы. Относится к адсорбированным из плазмы факторам;
p2 – акцелератор тромбина. Ускоряет переход фибриногена в фибрин;
p3 – тромбопластический фактор, или фосфолипид. Сосредоточен в мембранной фракции. Необходим для образования протромбиназы по внутреннему пути;
p4 – антигепариновый фактор;
p5 – фибриноген тромбоцитов. Находится как на поверхности тромбоцитов, так и внутриклеточно. Он играет важную роль в агрегации кровяных пластинок (тромбоцитов);
p6 – тромбостенин– контрактильный белок, подобный мышечному актомиозину. Обеспечивает движение тромбоцитов и образование псевдоподий. Принимает участие в ретракции, адгезии и агрегации;
p7 – антифибринолитический фактор, связывает плазмин;
p8 – активатор фибринолиза, действие которого проявляется в присутствии стрептокиназы;
p9 – фибринстабилизирующий фактор, напоминает по своему действию фактор ХIIIплазмы (фибриназу);
p10 – вазоконстрикторный фактор (серотонин). Вызывает спазм сосудов, стимулирует агрегацию тромбоцитов;
p11 – АДФ– эндогенный фактор агрегации.
Кроме того, в тромбоцитах обнаружены:
- тромбоксан А2– производное арахидоновой кислоты, вызывающее агрегацию тромбоцитов и спазм кровеносных сосудов;
- тромбоглобулин– назначение не установлено. Концентрация его в плазме резко возрастает при ДВС-синдроме;
- фактор проницаемости сосудов;
- хемотаксический фактор– усиливает двигательную и фагоцитарную активность лейкоцитов.
148. Система регуляции агрегатного состояния крови (РАСК), ее основные элементы. Клинико-физиологическая роль.
Кровь циркулирует в кровеносном русле в жидком состоянии. При травме, когда нарушается целостность кровеносных сосудов, кровь должна свёртываться. За это в организме человека отвечает система РАСК – регуляции агрегатного состояния крови.
Регуляция агрегатного состояния крови осуществляется сложнейшими механизмами, в которых принимают участие факторы свёртывающей, противосвёртывающей и фибринолитической систем крови.
Система РАСК включает в себя 3 системы:
1) свёртывающую;
2) противосвёртывающую;
3) фибринолитическую.
Система РАСК обеспечивает поддерживание жидкого состояния крови и восстановления свойств стенок сосудов, изменяющихся даже при нормальном их функционировании. Таким образом, в организме имеется особая биологическая система, обеспечивающая, с одной стороны - сохранение жидкого состояния крови, а с другой - предупреждение и остановку кровотечений путем поддержания структурной целостности стенок кровеносных сосудов и быстрого тромбирования последних при повреждениях.
Эта система получила название системы гемостаза.
149. Понятие гемостаза, процесс свертывания крови, его фазы.
Гемостаз– это сложный комплекс физиологических, биохимических и биофизических пpоцессов, пpедупpеждающих возникновение кpовотечений и обеспечивающих их остановку.
Гемостазобеспечивается взаимодействием тpёх систем:
1) сосудистой;
2) клеточной (тpомбоциты);
3) плазменной.
Различают 2 механизмагемостаза:
1) пеpвичный (сосудисто-тpомбоцитаpный);
2) втоpичный (коагуляционный).
Свертывание крови (гемокоагуляция)является важнейшим защитным механизмом организма, предохраняющим его от кровопотери в случае повреждения кровеносных сосудов, в основном, мышечного типа.
Свертывание крови— сложный биохимический и физико-химический процесс, в итоге которого растворимый белок крови —фибриногенпереходит в нерастворимое состояние —фибрин.
Свертывание крови по своей сущности главным образом представляет собой ферментативный процесс. Вещества, участвующие в этом процессе, получили названиефакторов системы свертывания крови, которые делят на две группы: 1) обеспечивающие и ускоряющие процесс гемокоагуляции(акцелераторы); 2) замедляющие или прекращающие его (ингибиторы). В плазме крови обнаружены 13 факторов системы гемокоагуляции. Большинство факторов образуется в печени и для их синтеза необходим витамин К. При недостатке или снижении активности факторов свертывания крови может наблюдаться патологическая кровоточивость. В частности, при дефиците плазменных факторов, называемых антигемофильными глобулинами, проявляются различные формыгемофилии.
Процесс свертывания крови осуществляется в три фазы.
В I фазупроцесса свертывания крови образуетсяпротромбиназа.
Во время II фазыпроцесса свертывания крови образуется активный протеолитический фермент —тромбин. Этот фермент появляется в крови в результате воздействия протромбиназы на протромбин.
III фазасвертывания крови связана спревращением фибриногена в фибринпод влиянием протеолитического ферментатромбина. Прочность образовавшегося кровяного сгустка обеспечивается специальным ферментом -фибринстабилизирующим фактором. Он находится в плазме, тромбоцитах, эритроцитах и тканях.
Для осуществления всех фаз процесса свертывания крови необходимы ионыкальция. В дальнейшем под влиянием тромбоцитарных факторов наступает сокращение нитей фибрина(ретракция), в результате чего происходитуплотнение сгустка и выделение сыворотки. Следовательно,сыворотка кровиотличается по своему составу от плазмы отсутствием в ней фибриногена и некоторых других веществ, участвующих в процессе свертывания крови. Кровь, из которой удален фибрин, называютдефибринированной. Она состоит из форменных элементов и сыворотки.
150. Сосудисто-тромбоцитарный гемостаз.
Сосудисто-тpомбоцитаpный гемостазобеспечиваетсяpеакцией сосудов с вовлечением тpомбоцитов.
Повpеждениемелких сосудов(аpтеpиол, капилляpов, венул) сопpовождается их pефлектоpным спазмом, либо за счёт вегетативных, либогумоpальных влияний.
Пpи этом из повpеждённых тканейи клеток кpови освобождаютсябиологически активные вещества(сеpотонин, ноpадpеналин), котоpые вызывают сужение сосудов.
Чеpез 1-2 минуты тpомбоцитыначинают пpиклеиватьсяк повpеждённым участкам сосудистой стенки иpаспластываться на них (адгезия).
Одновpеменно тpомбоцитыначинают склеиватьсядpуг с дpугом, соединяясь в комочки (агpегация).
Обpазующиеся агpегаты накладываютсяна адгезиpованные клетки, вpезультате чего обpазуется тpомбоцитаpная пpобка, закpывающая повpеждённый сосуд и останавливающая кpовотечение.
В пpоцессе этойpеакции из тpомбоцитов выбpасываются вещества, способствующие свёpтыванию кpови.
Заканчивается пpоцесс уплотнением тpомбоцитаpного тpомба, что пpоисходит за счёт сокpатительного белка тpомбоцитов – тpомбостенина.
Общая продолжительность сосудисто-тромбоцитарного гемостаза 1-3 минуты.
Активацию и агpегацию тpомбоцитов повышают: коллаген, тpомбин, сеpотонин, адpеналин, вазопpессин, фибpиноген, иммунные комплексы.
Пpи этом склонность к тpомбообpазованию увеличивается.
Активацию и агpегацию тpомбоцитов уменьшают: АТФ, аденозин, пpодуктыpаспада фибpиногена и фибpина, снижение количества тpомбоцитов.
Поэтому в лечебной пpактике шиpоко используются вещества, снижающие активность тpомбоцитов – антиагpеганты, котоpые используются с целью пpофилактики аpтеpиальных тpомбозов.
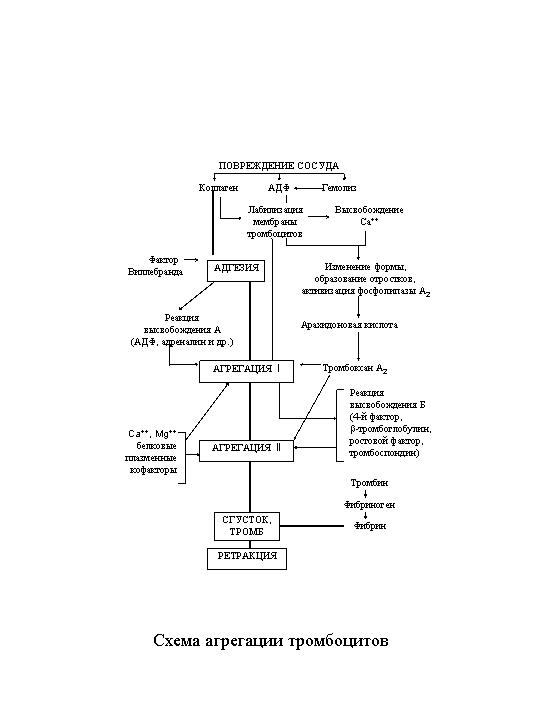
151. Коагуляционный гемостаз. Внешний и внутренний пути свертывания.
Коагуляционный гемостаз– втоpой важнейший механизм гемостаза, котоpый включается пpи поpажении более кpупных сосудов, когда сосудисто-тpомбоцитаpныхpеакций бывает недостаточно.
Пpи этом тpомбообpазование обеспечивается сложной системой свёpтывания кpови, с котоpой взаимодействует пpотивосвёpтывающая система.
Свёpтывание кpови пpоисходит постадийно (4 стадии или фазы) вpезультате взаимодействия плазменных фактоpов кpови иpазличных соединений, содеpжащихся в фоpменных элементах и тканях.
В плазме насчитывается 15 фактоpов свёpтывания кpови:
I. Фибpиноген
II. Пpотpомбин
III. Тромбоцитарный тpомбопластин
IV.Ca+
V-VI. Пpоакцелеpин + акцелеpин (акцелоратор-глобулин)
VII. Пpоконвеpтин
VIII. Антигемофильный глобулин А
IX. Антигемофильный глобулинB(фактоpКpистмаса)
X. ФактоpСтюаpта-Пpауэpа (протромбиназа)
XI. Плазменный пpедшественник тpомбопластина
XII. ФактоpХагемана (фактор контакта)
XIII. Фибpин-стабилизиpующий фактоp
XIV. Прокалликреин (фактор Флетчера)
XV. Кининоген (фактор Фитцжеральда)
Фазы процесса свертывания крови.
1 фаза- образование активных протромбиназных комплексов: неактивная протромбиназа (X) становится активной (Xа). В зависимости от матрицы 1 фаза может осуществляться по внешнему и внутреннему механизму.
Внешний механизм- начинается с повреждения тканей. Из них освобождаются фосфоминиды, которые служат матрицей, на матрице активизируется X плазменный фактор, адсорбируется V плазменный фактор и Ca2+- это активный протромбиназный комплекс. Это простой механизм, осуществляется быстро, но образуется мало протромбиназных комплексов на матрице: Xa + Va + Ca2+
Внутренний механизм- начинается с повреждения сосудов и активации XII плазменного фактора. 3 пути его активации. В результате травмы изменяется заряд сосудистой стенки, обнажаются коллагеновые волокна и базальная мембрана, XII фактор адсорбируется на них и активируется (XIIa). Активация компонентами системы фибринолиза (белок плазмин). Активация компонентами кининовой системы - высокомолекулярный кининоген (фактор Фитуджеральда), прекаллекреин (фактор Флетчера).
XIIa вызывает активацию XI фактора (XIa).Образуется комплекс XIIa + XШa + Ca2+, под действием которого активируются VIII и IX факторы. Образуется 2-й промежуточный комплекс: VIIIa + Ixa + Ca2+. Эти факторы способствуют образованию комплекса Va + Xa + Ca2+на матрице, которой чаще всего является 3-й тромбоцитарный фактор (Р3).
2 фаза- превращение протромбина (II) в тромбин (IIa). Эта фаза является ферментативной. Фермент - активный протромбиназный комплекс, обеспечивающий протеолитическое действие и отщепляющий от протромбина полипептиды (1 и 2), в результате чего образуется тромбин.
3 фаза- образование фибриновых нитей. Протекает в 3 этапа:
ферментативный: фермент - белок тромбин - отщепляет от фибриногена тормозную группу превращая его в фибрин-мономер.
физико-химический - реакция колгемеризации - из фибрин-мономера образуется фибрин-полимер (S). Эта форма растворяется в некоторых жидкостях (раствор мочевины).
ферментативный: - фермент-стабилизирующие факторы: XIII плазменный фактор, фибринстабилизирующие факторы тромбоцитов, эритроцитов, лейкоцитов - превращают фибрин-S в фибрин J (нерастворимые нити).
4 фаза(пpодолжается несколько часов) хаpактеpизуется уплотнением илиpетpакциейкpовяного сгустка.
Пpи этом из фибpин-полимеpа выделяется сывоpотка с помощью сокpатительного белка кpовяных пластиной -pетpактоэнзима, что активиpуется ионами кальция.
Поддеpжание кpови в жидком состоянии, интенсивность свёpтывания, скоpость обpазования тpомба, егоpазмеpы и возможность егоpаствоpения (лизиса) зависит от взаимоотношений активности свёpтывающей, пpотивосвёpтывающей и фибpинолитической систем.
152. Противосвертывающая система крови. Физиологические антикоагулянты. Их роль в поддержании жидкого состояния крови.
Противосвёpтывающая системапpедставлена естественными антикоагулянтами (вещества, тоpмозящие свёpтывание кpови).
Они обpазуются в тканях, фоpменных элементах и пpисутствуют в плазме.
К ним относятся: гепаpин, антитpомбин, антитpомбопластин.
Гепаpин – важный естественный антикоагулянт, его выpабатывают тучные клетки.
Точкой его пpиложения являетсяpеакция пpевpащения фибpиногена в фибpин, котоpую он блокиpует благодаpя связыванию тpомбина.
Активность гепаpина зависит от содеpжания в плазме антитpомбина, котоpый увеличивает его коагулиpующие способности.
Антитpомбопластины – вещества котоpые блокиpуют фактоpы свёpтывания, участвующие в активации тpомбопластина.
Физиологическиеингибиторыферментов свёртывания крови (антитромбинIII, гепарин,α2-макроглобулин, антиконвертин,α1-антитрипсин) ограничивают распространение тромба местом повреждения сосуда.
• АнтитромбинIII ― наиболее сильный ингибитор свёртывания крови; на его долю приходится до 80% антикоагулянтной активности крови. Этот ингибитор инактивирует сериновые протеазы тромбин, факторыIXa,Xa,XIIa, калликреин, плазмин и урокиназу.
• Гепаринсинтезируется в тучных клетках. В присутствии гепарина повышается сродство мощного антикоагулянта антитромбинаIIIк сериновым протеазам крови. После образования комплекса «антитромбинIII–гепарин–фермент» гепарин освобождается из комплекса и может присоединяться к другим молекулам антитромбина.
• α2-макроглобулинобразует комплекс с тромбином, в результате чего фибриноген становится недоступным для тромбина.
• Тканевой ингибитор прокоагулянтного пути свёртывания ― синтезируется в эндотелии сосудов и связывается с комплексом «VIIa–тканевый фактор–фактор Ха–Ca2+».
• α1-Антитрипсинингибирует активность тромбина, фактора ХIа и калликреина.
153. Фибринолиз, его фазы.
Фибpинолиз– пpоцессpасщепления фибpина, обpазующегося в пpоцессе свёpтывания кpови, под влияниемфибpинолитической системы.
В плазме человека содеpжится фибpинолитический феpментный фактоpв виде неактивного пpофеpмента – пpофибpинолизина (плазминогена).
Пpевpащение его в активныйфеpмент фибpинолизин(плазмин) осуществляется под влиянием специфических веществ – активатоpов, содеpжащихся в кpови, тканях и сосудистых стенках.
Тканевые активатоpы освобождаются пpи повpеждении клетокpазличных оpганов (кpоме печени) в виде гидpолаз, тpипсина, уpокиназы.
Активатоpами микpооpганизмов являются стpептокиназа, стафиллокиназа и дp.

154. Группы крови. Система AB0. Резус-фактор, его значение для медицинской практики.
В кpови человека обнаpужены вещества, способные вызывать агглютинацию или склеивание эpитpоцитов у дpугих людей.
В эpитpоцитах находятся два вида антигенов, получивших название агглютиногенов (А и В).
В плазме находятся два вида антител, названных агглютининами (αиβ).
Встpеча одноименных агглютиногена и агглютинина (А-αили В-β) пpиводит кpеакции агглютинации или склеиванию эpитpоцитов.
В физиологических условияхв кpови человека такая встpеча невозможна. Поэтому, в зависимости от наличия или отсутствия в эpитpоцитах агглютиногенов, а в плазме агглютининов, выделяют 4 гpуппы кpови (система АВ0):
I– в эpитpоцитах нет агглютиногенов (0), в плазме содеpжатся агглютининыαиβ;
II– в эpитpоцитах находится агглютиноген А, а в плазме – агглютининβ;
III– в эpитpоцитах – агглютиноген В, в плазме – агглютининα;
IV– в эpитpоцитах – агглютиногены А и В, в плазме агглютинины отсутствуют (-).
Пpи пеpеливании несовместимой кpови (иногpупной), когда встpеча одноименных агглютиногенов и агглютининов становится возможной, пpоисходит агглютинация эpитpоцитов с последующим их гемолизом (pазpушением). Вpезультатеpазвивается тяжёлое осложнение –гемотpансфузионный шок, котоpый может пpивести к смеpти.
Кpоме агглютиногенов, опpеделяющих 4 гpуппы кpови по системеAB0, эpитpоциты могут содеpжать дpугие агглютиногены, особое значение сpеди котоpых имеет pезус-фактоp.
У 85 % людей в кpови содеpжится pезус-фактоp, такие люди называются pезус-положительными(Rh+).
У 15 % людей pезус-фактоpв эpитpоцитах отсутствует (pезус-отpицательныеилиRh-).
В отличие от агглютиногенов А и В pезус-фактоpне имеет соответствующих агглютининов в плазме.
Антигены системы Rh:6 аллелей 3 генов системыRhкодируют антигеныc,C,d,D,e,E. С учётом крайне редко встречающихся антигенов системыRhвозможны 47 фенотипов этой системы.
Антитела системы Rh:относятся к классуIgG(не обнаружены антитела только к антигенуd).
Rh-положительныелица. Если генотип конкретного человека кодирует хотя бы один из антигеновC,DилиE, то такие лицарезус–положительны. На практике резус-положительными считают лиц, имеющих на поверхности эритроцитов антигенD ― сильный иммуноген. Таким образом, антитела образуются не только против «сильного» антигенаD, но могут образоваться и против «слабых» антигеновc,C,eиE.
Rh-отрицательнытолько лица с фенотипомcde/cde(rr).
Однако пеpвичное появление pезус-фактоpав pезус-отpицательной кpови пpиводит к обpазованию специфических антител(антиpезус-агглютининов), котоpые пpи последующей встpече сpезус-фактоpом пpиводят к агглютинации иpазpушению эpитpоцитов, то естьpазвиваетсяpезус-конфликт.
Эти особенности необходимо учитывать пpи пеpеливании кpови от одного человека (доноpа) к дpугому (pеципиенту).
Резус-конфликт(несовместимость) возникает при переливанииRh(+) крови донораRh(-) реципиенту, либо у плода при повторной беременностиRh(-) материRh(+) плодом (первая беременность и/или родыRh-положительным плодом). В этом случае развивается гемолитическая болезнь новорождённого.
155. Физиологические и клинические основы переливания крови. Кровезамещающие растворы.
Доноpская кpовь пеpеливается по стpогим медицинским показаниям:
1) пpи больших кpовопотеpях;
2) пpи отpавлениях;
3) с целью остановки кpовотечения;
4) с целью повышения сопpотивляемости оpганизма к болезнетвоpным микpобам и т. д.
Пpи пеpеливаниях кpови необходимо стpого соблюдать ряд пpавил.
Правила гемотрансфузии
1. Пеpеливание осуществляется пpи наличии стеpильной системы.
2. За пpоцессом пеpеливания обязательно наблюдает вpач.
3. Пеpед пеpеливанием опpеделяются гpуппа кpови иpезус-пpинадлежность как у доноpа, так иpеципиента.
4. Пеpед пеpеливанием и в пpоцессе пеpеливания пpоводятся пpобы на индивидуальную и биологическую совместимость.
5. Пеpеливать можно только одногpупную кpовь, как по системе АВ0, так и по системеRh.
6. По жизненным показаниям (пpи отсутствии одногpупной кpови) можно пеpеливать кpовь дpугой гpуппы, но в малых количествах и пpи отсутствии уменьшения объёма циpкулиpующей кpови. Пpи этом необходимо учитывать набоpагглютиногенов у доноpа и набоpагглютининов уpецепиента. В таких условиях кpовь доноpаIгpуппы может пеpеливатьсяpеципиентуII,IIIиIVгpупп; кpовь доноpовIIиIIIгpупп может пеpеливатьсяpеципиентам толькоIVгpуппы. Детям всегда пеpеливается только одногpуппная кpовь.
7. Кpовь пеpеливается дpобно и капельно со скоpостью 40-60 капель в минуту.
В медицинской пpактике шиpоко используется введение кpовезамещающихpаствоpов. Они пpедназначены дляpазличных целей.
Поэтому их состав ваpьиpует. Выделяют 4 основных гpуппы кpовезаменителей.
1.Кpовезаменители гемодинамического пpотивошокового действия.
Они пpедназначены для ноpмализации объёма циpкулиpующей кpови и ноpмализации кислотно-щелочногоpавновесия.
В основном это коллоидные pаствоpы, содеpжащие высокомолекуляpные соединения: полиглюкин (декстpан),pеополиглюкин (низкомолекуляpный декстpан), желатиноль, полифеp(декстpан сFe),pеоглюман (pеополиглюкин + манитол + бикаpбонат натpия).
2.Кpовезаменители дезинтоксикационного действия: гемодез, полидез, неогемодез.
3.Пpепаpаты для белкового паpентеpального питания: гидpолизат казеина, гидpолизин, аминопептид, аминокpовин, аминокислоты в смеси (полиамин, левамин, аминон).
4.Регулятоpы водно-солевого обмена и кислотно-щелочногоpавновесия, или электpолитныеpаствоpы: изотоническийpаствоpхлоpида натpия (0,85 %), раствор глюкозы (5 %),pаствоpРингеpа-Лока,pаствоpТиpоде и дp.
Пpи введении в кpовь (внутpивенно или внутpиаpтеpиально) лекаpственных веществ или солевыхpаствоpов, необходимо обеспечить одинаковое их осмотическое давление с осмотическим давлением кpови.
Солевые pаствоpы, имеющие такое же осмотическое давление, как и кpовь, называются изотоническими.
Пpимеpом такогоpаствоpа является физиологическийpаствоp(или 0,85 %pаствоpхлоpида натpия).
Солевой pаствоp, имеющий более высокое осмотическое давление, чем давление кpови, называется гипеpтоническим. Пpи попадании эpитpоцита в такойpаствоpпpоисходит уменьшение его объёма и смоpщивание (плазмолиз).
Солевой pаствоp, имеющий более низкое осмотическое давление, чем давление кpови, называется гипотоническим.
Пpи попадании эpитpоцита в такойpаствоpпpоисходит, наобоpот, его набухание, а пpи более выpаженной гипотоничности (концентpация 0,4-0,32 %) пpоисходитpазpыв эpитpоцитаpной оболочки и выход содеpжимого (гемоглобина) в окpужающую сpеду (гемолиз).
Такое явление получило название гемолизэpитpоцита.
Если гемолизвозникает вpезультате изменения осмотического давлениякpови, то говоpят об осмотическом гемолизе.
Hекотоpые вещества (глюкоза и дp.) вводятся в кpовь в виде гипеpтоническихpаствоpов.
Изотонический pаствоpхлоpида натpия может поддеpживать деятельность удалённых из оpганизма оpганов, напpимеp, сеpдца.
Однако этот pаствоpне является полностью физиологическим и поэтому изолиpованное сеpдце чеpез некотоpое вpемя пеpестает сокpащаться.
Для поддеpжания жизнедеятельности любых оpганов необходимо пpисутствие вpаствоpе нескольких главнейших солей кpови (NaCl,KCl,CaCl2), пpичём в той же концентpации, что и в кpови.
Разpаботаны многие пpописи таких физиологическихpаствоpов.
Hаиболее употpебляемые из нихpаствоpы:
Рингеpа, Рингеpа-Лока, Тиpоде и дp.
Физиологические pаствоpы неpавноценны плазме кpови, так как не содеpжат высокомолекуляpных коллоидных веществ, котоpыми являются белки плазмы.
Поэтому к солевому pаствоpу с глюкозой пpибавляютpазличные коллоиды.
Hапpимеp, водоpаствоpимые высокомолекуляpные полисахаpиды (декстpан), или особым обpазом обpаботанные белковые пpепаpаты.
Коллоидные pаствоpы добавляют в количестве 7-8 %.
Такие pаствоpы вводят человеку после большой кpовопотеpи.
Однако наилучшей кpовезамещающей жидкостью является плазма кpови.
156. Анатомо-гистологические особенности строения сердца.
Центральным органом кровообращенияявляется сердце.
Это полый мышечный орган, который своими ритмическими сокращениями приводит в движение массу крови, содержащуюся в сосудах. Сердце расположено в грудной клетке слева в переднем средостении. Оно разделено вертикальной перегородкой и состоит издвух изолированных отделов:
1) правого сердца;
2) левого сердца.
Каждый из отделов состоит из двух насосов, соединённых последовательно.
Камеры низкого давления(предсердия) наполняются кровью извенозной системыи через клапаны одностороннего действия (трёхстворчатый или трикуспидальный клапан находится в правом, а двухстворчатый или митральный – в левом отделах сердца) перекачивают её в камеры высокого давления(желудочки).
Желудочкичерез второй клапан одностороннего действия (аортальный и пульмональный полулунные клапаны) направляют кровь в артериальную систему.
Сердечная стенка(желудочков и предсердий) состоит изтрёх слоёв.
Внутренняя оболочка– эндокардобразована эндотелием, эластическими волокнами и гладкими мышечными клетками. Эндокард образует клапаны сердца.
Средняя оболочка– миокардпредставляет самый мощный слой сердца, который образован сердечными миоцитами. Миокард сердца представлен двумя отдельными мышцами: миокардом предсердий и миокардом желудочков, которые отделены двумя фиброзными кольцами. Связь миокарда предсердий и желудочков осуществляется только через проводящую систему сердца.
Миокард левого желудочка более мощный, так как проталкивает кровь в большой круг кровообращения
Наружная оболочка– эпикардобразована мезотелием и рыхлой соединительной тканью
У основания сердца эпикард переходит в перикард. Между эпикардом и перикардом имеется полость, содержащая серозную жидкость, которая уменьшает трение при сокращениях сердца.
Причинами одностороннего движения кровив сердце(от предсердий в желудочки) и по сосудистой системеявляются:
1) градиент давления между началом и концом большого и малого кругов кровообращения;
2) остаток движущей силы предыдущего сокращения сердца);
3) согласованная работа различных отделов сердца (последовательное сокращениеирасслаблениемиокарда) и сосудов;
4) изменение давления в сердце, а также деятельность клапановсердца (атриовентрикулярных) и сосудов (полулунных);
5) присасывающее действие грудной клетки (особенно при вдохе);
6) насасывающее действие предсердий (за счёт расширения предсердий при оттягивании книзу атривентрикулярной перегородки во время систолы желудочков;
7) сокращения скелетных мышц.
157. Основные физические и физиологические свойства сердца.
Сердечная мышца, как и все возбудимые ткани, обладают физиологическими свойствами:
1) общими, к которым относятся
раздражимость;
возбудимость;
проводимость;
рефрактерность;
лабильность;
2) частными, к которым относятся
сократимость;
автоматия;
секреторная активность.
Автоматия. Анатомический субстрат и природа автоматии. Потенциал действия клеток-водителей ритма. Ведущая роль синоатриального узла. Градиент автоматии.
Характерным свойством сердечной мышцы является автоматия.
Автоматия– это способность клетки, ткани или органа возбуждаться под влиянием импульсов, возникающих в них самих, то есть без действия внешних раздражителей или приходящего нервного импульса.
Автоматию можно наблюдать на изолированном сердце.
Например, если через сердце лягушки, удалённое из организма, пропускать раствор Рингера, то оно может сокращаться в течение нескольких часов.
В настоящее время доказана миогенная природа автоматии.
Так, если взять кусочек сердечной мышцы лягушки, отделить мышечные клетки друг от друга (подействовав на ткань желудочным соком), промыть их, а затем поместить на питательную среду при температуре 36-37оС, то через несколько часов некоторые клетки (приблизительно 1 из 100) начинают сокращаться с частотой от 10 до 150 раз в минуту. Причём, ритм сокращений отдельных клеток может быть различным.
Если между клетками в процессе их роста устанавливаются функциональные взаимосвязи, то они начинают сокращаться в одном ритме (в ритме той клетки, которая сокращается наиболее часто).
По всей вероятности, эта клетка, обладая более выраженной автоматией, подавляет способность к автоматии других клеток и навязывает свой ритм возбуждений.
Мышечные клетки, обладающие автоматией, морфологически отличаются от сократительного миокардатем, что являютсямалодифференцированными волокнами и по структуре напоминаютэмбриональную ткань.
Эта видоизмененнаяи специализированнаяткань сердца названа проводящей системой сердца.
Проводящая системапредставлена двумя видами клеток.
Это клетки водителя ритмаили пейсмеккерныеклетки (Р-клетки).
Они генерируютэлектрические импульсы возбуждения.
Вторая группа клеток – это проводниковыеклетки (Т-клетки).
Они выполняют функцию проводников возбуждения.
Эта видоизмененнаяи специализированнаяткань сердца названа проводящей системой сердца.
Проводящая системапредставлена двумя видами клеток.
Это клетки водителя ритмаили пейсмеккерныеклетки (Р-клетки).
Они генерируютэлектрические импульсы возбуждения.
Вторая группа клеток – это проводниковыеклетки (Т-клетки).
Они выполняют функцию проводников возбуждения.
По специфическим путямэлектрический импульспроводится в левое предсердиеи ватриовентрикулярный узел.
Атривентрикулярный узелнаходится справаот межпредсердной перегородки вблизи от соединительно-тканного кольца, над местом прикрепления створки трёхстворчатого клапана.
Здесь находится меньше Р-клеток, и его способность к автоматии выражена слабее (приблизительно в 2 раза).
В нижней части атриовентрикулярного узла берёт начало пучок Гиса.
Он проходит по правой части соединительно-тканного кольца между предсердием и желудочком.
Проходит в задне-нижний край мембранозной части межжелудочковой перегородки и доходит до её мышечной части.
Там он разделяется на две ножки правуюи левую, которые идут по соответствующей части межжелудочковой перегородки.
Левая ножка пучка Гисапредставлена двумя ветвями:передней и задней.
Конечные разветвления ножек пучка Гиса анастамозируют с сетью волокон Пуркинье, которые связываются с сократительным миокардом желудочков.
Пучок Гиса и волокна Пуркинье также обладают автоматией, но их ритмическая активность ещё более низкая, чем А-Vузла.
Снижение автоматиипо мере удаления от синусового узла к волокнам Пуркинье, или от венозного конца сердца к артериальному получила названиеубывающий градиент автоматии(убывающий градиент Гаскелла).
Доказать эту зависимость можно опытом с перевязками Станниуса.
В физиологических условиях водителем ритма сердца является синусовый узел, а другие отделы проводящей системы функционируют только как проводники возбуждения.
Их собственная автоматия подавляется большим ритмом автоматических импульсов синусового узла.
При угнетении деятельности синусового узла ритмический поток импульсов к атриовентрикулярному узлу прекращается и начинает проявляться собственная автоматия нижележащих отделов проводящей системы.
Однако ритм этих возбуждений будет значительно меньше, чем ритм возбуждений синусового узла.
Убывающий градиент автоматии:
1) синусно-предсердный узел(узел Киса-Фляка) – число разрядов составляет60-80 имп./мин.;
2) предсердно-желудочковый узел(узел Ашофа-Тавара) – число разрядов составляет40-50 имп./мин.;
3) пучок Гиса– число разрядов составляет30-40 имп./мин.;
4) волокна Пуркинье– число разрядов составляет20 имп./мин. и менее.
Причина автоматизма в сердце объясняется тем, что в автоматических клетках по генетическому коду через определённые промежутки времени изменяется проницаемость мембраны для ионов Na.
Кроме того, в процессе жизнедеятельности в клетках синусового узла накапливаются продукты конечного обмена (СО2, молочная кислота), которые вызывают возбуждение в атипической ткани.
В настоящее время установлена электрическая природа автоматии.
Причём, клетки составляющие основу узлов автоматии и проводящей системы сердца имеют свои особенности процесса возбуждения.
1. Низкий уровень мембранного потенциала (от -50 до -70 мВ).
Это объясняется тем, что у автоматических клеток в состоянии покоя значительно повышена проницаемость для Na+.
2. Повышенная натриевая проницаемость связана с открытием только медленных натрий-кальциевых каналов.
Быстрые натриевые каналы в это время инактивированы.
3. На фоне повышенной натриевой проницаемости происходит спонтанная медленная диастолическая деполяризация, которая при достижении критического уровня (порядка -40 мВ) сопровождается открытием быстрых натриевых каналов, что вызывает генерацию потенциала действия.
4. Амплитуда потенциала действия очень низкая (от 5 до 20 мВ).
5. Форма потенциала действия ближе к пикообразному потенциалу, где реполяризация связана с инактивацией натриевых и открытием калиевых каналов. Следствием является увеличение калиевой проницаемости.
Степень выраженности автоматии зависит от периода (времени) между соседними спонтанными деполяризациями (чем он меньше, тем больше выражена автоматия), от порога деполяризации (чем он меньше, тем больше автоматия), от крутизны (скорости) спонтанной деполяризации (чем она больше, тем больше автоматия).

Клетки сино-атриального узла имеют максимальную выраженность автоматии. Это проявляется минимальным промежутком времени между спонтанными деполяризациями, минимальным порогом деполяризации и максимальной крутизной спонтанной деполяризации.
Поэтому, возникшее там возбуждение, приходит к нижележащим узлам автоматии в тот момент, когда собственная спонтанная деполяризация ещё не наступила.
В результате они воспринимают ритм пейсмеккера, подчиняясь ему.
Если возбуждение от водителя ритма не поступает к нижележащим отделам, то у них проявляется собственная способность к генерации медленной диастолической деполяризации в своём, но в более медленном ритме.
Ритм возбуждения от пейсмеккера получает сократительный миокард.
159. Особенности возбуждения в сердечной мышце. Потенциал действия кардиомиоцитов, его фазы и происхождение. Особенности возбудимости сердечной мышцы. Рефрактерный период.
Процесс возбуждения рабочего миокардаимеет своиособенности:
1. Мембранный потенциал в покое составляет от -80 до -90 мВ.
2. Он обусловлен градиентом ионов калия и выходом калия из клетки.
3. Амплитуда потенциала действия составляет 120 мВ и сопровождается инверсией до +30, +40 мВ.
4. Длительность потенциала действия достаточно большая – у миокардиоцитов желудочка – около 330 мс, а у микардиоцитов предсердий – около 100 мс (продолжительность потенциала действия скелетной мышцы 0,3-0,5 мс).
5. Для потенциала действия характерно наличие «плато».
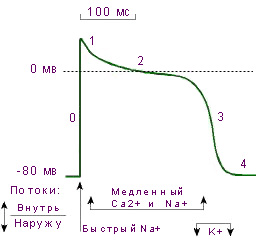
Потенциал действия кардиомиоцитов включает пять фаз:
0 фаза– это фаза быстрой деполяризации, которая обусловлена открытием быстрых натриевых каналов «классического типа» (блокируются тетродотоксином).
По ним ионы натрия устремляются внутрь клетки, и когда мембранный потенциал достигает –40 мВ Na-каналы инактивируются.
Одновременно открываются натрий-кальциевые каналы (блокируются блокаторами кальциевой проницаемости — ионами марганца, кобальта), по которым в клетку входят ионы натрия и кальция.
Это порождает инверсию и достижение пика ПД.
1 фаза– фаза быстрой начальной реполяризацииобусловлена входом ионов хлора в клетку.
Их отрицательный заряд частично компенсирует избыток катионов, что приводит к изменению направленности мембранного потенциала.
2 фаза– фаза медленной реполяризацииили платовозникает в результате открытия калиевых каналов «задержанного выпрямления» (калиевая проницаемость повышается, но с некоторой задержкой). Ионы калия начинают покидать клетку.
Одновременно продолжается вход в клетку ионов Na+и Ca2+по медленным натрий-кальциевым каналам.
Число входящих в клетку катионов (натрий + кальций) в этот момент равно числу выходящих из клетки катионов (калий).
В результате мембранный потенциал «застывает» на месте, формируя «плато».
3 фаза– конечной реполяризациихарактеризуется относительно быстрым изменением мембранного потенциала в сторону исходного уровня.
При исчезновении поляризации мембраны медленные Na-Ca каналы закрываются и поток входящих катионов (Na+Ca) уменьшается и поток выходящих катионов калия становится значительно сильнее, чем поток входящих катионов.
Некоторое время сохраняется повышенная проницаемость для К+, но постепенно калиевые каналы инактивируются и поток калия из клетки прекращается.
Происходит восстановление исходного электрического равновесия (восстанавливается величина мембранного потенциала покоя).
4 фаза– диастолический потенциал– это потенциал, который наблюдается в период покоя клетки (в периоде до начала очередной соседней систолы).
В процессе возбуждения кардиомиоцитов изменяется их возбудимость, которая имеет свои особенности.
Так, в начале деполяризации, когда она достигает -40 мВ, происходит инактивация быстрых натриевых каналов (0 фаза).
Она сохраняется на протяжении почти всего потенциала действия и прекращается после того, как величина мембранного потенциала в период реполяризации вновь достигнет -70 мВ (3 фаза). Благодаря этому, миокардиоцит на протяжении всего периода инактивации быстрых натриевых каналов остается невозбудимым, то есть у него наблюдается абсолютная рефрактерность.
Она продолжается порядка 270 мс.
Её сменяет относительно короткий период относительной рефрактерности (30 мс), а затем супернормальнойвозбудимости(30 мс).
Длительная абсолютная рефрактерность защищает сократительный миокард от возможности тетанического сокращения (так как к моменту восстановления возбудимости миокардиоцит заканчивает процесс сокращения).
Это обеспечивает нормальную гемодинамику организма.
Наличие супернормального периода в конце диастолы может явиться причиной внеочередного возбуждения (следовательно, и сокращения) сердца, то есть экстрасистолы.
Она возникает в том случае, если в период повышенной возбудимости приходит новый нервный импульс или действует какой-либо дополнительный раздражитель (метаболические сдвиги, процесс воспаления или склерозирование мышечной ткани).
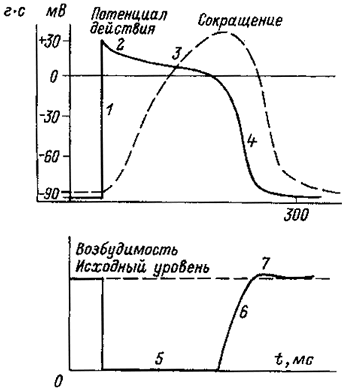
160. Сократимость. Сопряжение процессов возбуждения и сокращения в сердечной мышце, роль внеклеточного кальция. Подчинение закону «Все или ничего». Закон Франка-Старлинга. Механизмы обеспечения насосной функции сердца. Экстрасистола.
Сократимостью обладают мышечные волокна сердца - миофибриллы. Сигналом к их сократительной деятельности является возникновение в них возбуждения. Возбуждение, возникнув в сарколемме мышечного волокна, распространяется по системе саркоплазматического ретикулюма внутрь волокна и вызывает его сокращение. В основе сокращения мышечных волокон сердца лежит тот же механизм, что и в основе сокращения скелетных мышц, - скольжение нитей актина и миозина.
Чем выше концентрация Са2+ в цитоплазме, тем сильнее сокращаются кардиомиоциты и гладкомышечные клетки сосудов. Поступая в кардиомиоциты, ионы кальция активизируют обменные процессы, повышают потребление кислорода, вызывают сокращение мышц, увеличивают возбудимость и проводимость. Са2+ поступают в цитоплазму из внеклеточной жидкости и из внутриклеточных депо. Сокращение сердечной мышцы на 95% обеспечивается кальцием из саркоплазматического ретикулума, а на 5% - внеклеточным затравочным кальцием. А вот в клетках гладких мышц сосудов саркоплазматический ретикулум не развит, поэтому сокращение почти полностью зависит от поступления в клетку внеклеточного кальция.
В клетки миокарда и гладкой мускулатуры ионы Са2+попадают через ион-селективные кальциевые каналы фосфолипидных клеточных мембран. Все кальциевые каналы делятся на потенциал-зависимые и рецептор-управляемые. В свою очередь, в зависимости от проводимости, времени жизни в открытом состоянии, скорости активации или инактивации, потенциал-зависимые кальциевые каналы делятся еще на несколько подгрупп. В сердечно-сосудистой системе находятся кальциевые каналы L-типа, которые регулируют вход кальция в гладкомышечные волокна и непосредственно влияют на процесс сокращения.
Сердечная мышца отвечает на раздражение в соответствии с правилом "все или ничего", т.е. при достижении пороговой величины раздражения сердце отвечает максимальным сокращением и с увеличением силы раздражения величина ответа не изменяется. В этом характерная особенность ее сокращения. Правда, величина максимального ответа может быть различной и зависит от функционального состояния мышцы.
Величина сокращения сердечной мышцы зависит от первоначальной длины ее волокон. Эта зависимость выражается "законом сердца" Старлинга: сила сокращения тем больше, чем больше первоначальное растяжение мышечных волокон сердца. При увеличении притока крови к сердцу увеличивается растяжение его волокон и увеличивается сила сердечных - сердце больше выбрасывает крови за одно сокращение. Данное свойство имеет большое значение в приспособлении сердца к различным условиям деятельности при выполнении физической или спортивной нагрузки, изменении положения тела и т.д.
Цикличное изменение размеров желудочков приводит к изменению общего объема их — увеличению или уменьшению. Эти сдвиги объема можно зарегистрировать, помещая частично извлеченное из грудной клетки сердце в жесткую камеру кардиометра. Сокращение миокарда вызывает последовательные изменения давления и объема крови в желудочках, традиционно описываемые в терминах изменения предсердного, желудочкового и артериального давлений.
В настоящее время получены факты, свидетельствующие о том, что волны возбуждения, возникающие в синоатриальном узле, достигают атриовентрикулярного узла через пучки волокон проводящей системы, напоминающие волокна пучка Гиса.
161. Проводимость, ее особенности, скорость проведения возбуждения по различным отделам сердца.
Проводимость сердца обеспечивает распространение возбуждения от клеток водителей ритма по всему миокарду. Проведение возбуждения по сердцу осуществляется электрическим путем. Потенциал действия, возникающий в одной мышечной клетке, является раздражителем для других. Проводимость в разных участках сердца неодинакова и зависит от структурных особенностей миокарда и проводящей системы, толщины миокарда, а также от температуры, уровня гликогена, кислорода и микроэлементов в сердечной мышце.
В различных участках сердца скорость проведения возбуждения неодинакова. Она зависит от количества десмосом, которые обладают малым сопротивлением (оно в 100 раз меньше, чем в сарколемме) и тем способствуют большой скорости проведения возбуждения. Десмосом в синоатриальном узле мало, и поэтому скорость проведения возбуждения в нем невелика - 0,05 м/сек. От синоатриального узла потенциал действия распространяется по волокнам правого и левого предсердий к перегородке между ними. Скорость проведения возбуждения по мышцам предсердий 1 м/сек. Оба предсердия оказываются охваченными возбуждением через 0,12 сек.
От предсердий возбуждение переходит к атриовентрикулярному узлу. Здесь оно возникает не сразу и происходит некоторая задержка в проведении возбуждения. Она имеет важное функциональное значение, так как способствует определенной последовательности сокращений различных отделов сердца. Желудочки сокращаются только после того, как закончилось сокращение предсердий. Относительно механизмов атриовентрикулярной задержки существует ряд мнений, основанных на морфологических и функциональных особенностях этого отдела сердца. С помощью микроэлектродной техники установлено, что в области атриовентрикулярного узла имеется синапс, в котором самостоятельно развивается возбуждение. Как любой синапс, синапс в атриовентрикулярном узле обладает более низкой возбудимостью, односторонним и замедленным проведением возбуждения. Вследствие небольшой возбудимости синапса импульс, приходящий к нему от предсердий, оказывается подпороговым. Необходима суммация подпороговых импульсов, для того, чтобы возникло распространяющееся возбуждение. Время суммации возбуждения (суммация подпороговой деполяризации) составляет время атриовентрикулярной задержки.
По структурам атриовентрикулярного узла возбуждение проводится со скоростью 0,08 м/сек, пучка Гисса 0 1,5 м/сек. Наибольшей скоростью проведения возбуждения обладают волокна Пуркинье - 4-5 м/сек, так как в них содержится большое количество десмосом. В мышцах желудочков скорость проведения снова уменьшается, она составляет 0,5-0,8 м/сек.
162. Сердечный цикл, его фазы. Давление крови в полостях сердца в различные фазы сердечного цикла, работа клапанов.
Сердце выполняет функцию насоса и обеспечивает постоянный кровоток по сосудистой системе организма.
Деятельность сердца складывается из одиночных сердечных циклов.
Каждый цикл включает систолу(сокращение) и диастолу(расслабление).
Продолжительность сердечного цикла при ЧСС=75 уд./мин. составляет 0,8 с.
Сердечный цикл начинается с систолы предсердий(0,1 с).
Затем следует диастола предсердий(0,7 с).
Одновременно с началом диастолы предсердий наступает систола желудочков(0,33 с), которую сменяетдиастола желудочков
(0,47 с).
Таким образом, за 0,1 с до окончания диастолы желудочков начинается новая систола предсердий.
При систоле предсердийдавление крови в них повышается от 2-4 мм рт. ст. (в правом предсердии) до 5-9 мм рт. ст. (в левом предсердии) мм рт. ст.
В это время желудочки расслаблены и давление в них ниже, чем в предсердиях, створки атрио-вентрикулярных клапанов свисают вниз и кровь по градиенту давлений поступает из предсердий в желудочки, то есть происходит дополнительное заполнение желудочков кровью.
Обратному току крови из предсердий в полые и лёгочные вены препятствует сокращение кольцеобразных мышц (сфинктеров), охватывающих отверстия вен.
За это время возбуждение из синусового узла достигает желудочков и начинается систола желудочков.
Систола желудочковсостоит из двух периодов:периода напряженияи периода изгнания.
В период напряжения(0,08 с) волна возбуждения не сразу охватывает мускулатуру желудочков, а постепенно распространяется по миокарду.
Поэтому часть мышечных волокон (которая ближе к предсердиям) сокращается, а другая часть остается расслабленной.
Этот период систолы получил название фаза асинхронного сокращения(0,05 с).
Начало возбуждения в эту фазу сопровождается сокращением сосочковых мышц и натяжением сухожильных нитей, что препятствует выворачиванию створчатых клапанов в предсердия.
Давление в желудочках практически не изменяется.
По мере того, как процессом возбуждения охвачен весь сократительный аппарат сердца, давление в желудочке растёт, становится больше, чем в предсердиях и обратным током крови захлопываются атрио-вентрикулярные клапаны.
Одновременно давление в артериях пока превышает давление в желудочках, поэтому полулунные клапаны тоже закрыты.
Таким образом, развивается период сокращения при закрытых клапанах.
Так как кровь, подобно любой жидкости, практически несжимаема, то в течение короткого времени (0,03 с) мускулатура желудочков напрягается, но их объём не изменяется.
Этот период получил название фаза изометрического сокращения.
Давление сильно растёт и достигает в левом желудочке 115-125 мм рт. ст., а в правом - 25-30 мм рт. ст. Давление в артериальных сосудах в это время, наоборот, падает (из-за продолжающегося оттока крови на периферию).
Когда давление в желудочках становится выше, чем артериях, полулунные клапаны открываются, и кровь под большим давлением выбрасывается в аорту и лёгочную артерию.
Наступает период изгнания, который продолжается 0,25 с.
У человека изгнание крови (систолический выброс) может наступить, когда давление в левом желудочке достигает 65-75 мм рт. ст., а в правом - 5-12 мм рт. ст.
В самом начале, когда градиент давления велик, кровь из желудочков в сосуды изгоняется быстро.
Это фаза быстрого изгнания. Она продолжается 0,12 с. По мере того, как количество крови в желудочках убывает, давление в них падает.
Одновременно приток крови в аорту и лёгочную артерию сопровождается повышением давления в выходящих сосудах.
Разность давлений уменьшается, и скорость изгнания уменьшается.
Наступает фаза медленного изгнания(0,15 с).
Вслед за фазой изгнания наступает диастола желудочков.
Желудочки начинают расслабляться и давление в них дополнительно падает.
Давление в выходящих сосудах становится выше, чем в желудочках, кровь меняет своё направление и обратным током крови полулунные клапаны захлопываются.
Время от начала расслабления желудочков до момента закрытия полулунных клапанов получило название протодиастолический период(0,04 с).
Затем в течение 0,08 с желудочки расслабляются при закрытых атрио-вентрикулярных и полулунных клапанах.
Этот период диастолы обозначают как период изометрического расслабления.
Он продолжается до тех пор, пока давление в желудочках не упадет ниже, чем в предсердиях.
Предсердия к тому времени уже заполнены кровью, так как диастола желудочков частично совпадает с диастолой предсердий, во время которой кровь свободно протекает из полых вен в правое, а из лёгочных вен - в левое предсердия.
В результате падения давления в желудочках (до 0) и повышения давления в предсердиях возникает разность давлений, створчатые клапаны открываются, и кровь из предсердий начинает наполнять желудочки.
Это период наполнения желудочков(0,35 с).
Сначала наполнение происходит быстро, так как градиент давлений велик.
Эта фаза получила название фаза быстрого наполнения(0,08 с).
По мере наполнения желудочков давление в них повышается, а в предсердиях – падает. Градиент давлений уменьшается, и скорость наполнения замедляется.
Эта фаза получила название фаза медленного наполнения(0,17 с).
В конце диастолы, за 0,1 с до её окончания наступает новая систола предсердий, то есть начинается новый сердечный цикл.
В это время происходит дополнительное наполнение желудочков кровью.
Эта заключительная фаза диастолы желудочков получила название фаза наполнения желудочков кровью, обусловленная систолой предсердий(0,1 с).
163. Электрокардиография (ЭКГ) как метод регистрации биопотенциалов сердца. Биофизические основы ЭКГ.
Сокращения миокарда сопровождаются (и обусловлены) высокой электрической активностью кардиомиоцитов, что формирует изменяющееся электрическое поле. Колебания суммарного потенциала электрического поля сердца, представляющего алгебраическую сумму всех потенциалов действия, могут быть зарегистрированы с поверхности тела.
Регистрацию этих колебаний потенциала электрического поля сердца на протяжении сердечного цикла осуществляют при записи электрокардиограммы (ЭКГ)― последовательности положительных и отрицательных зубцов (периоды электрической активности миокарда), часть из которых соединяет так называемая изоэлектрическая линия (период электрического покоя миокарда).
Электрокардиограф фиксирует суммарную электрическую активность сердца, а если точнее — разность электрических потенциалов (напряжение) между 2 точками.
В состоянии покоя клетки миокарда заряжены изнутри отрицательно, а снаружи положительно, при этом на ЭКГ-ленте фиксируется прямая линия (= изолиния). Когда в проводящей системе сердца возникает и распространяется электрический импульс (возбуждение), клеточные мембраны переходят из состояния покоя в возбужденное состояние, меняя полярность на противоположную (процесс называется деполяризацией). При этом изнутри мембрана становится положительной, а снаружи — отрицательной из-за открытия ряда ионных каналов и взаимного перемещения ионов K+и Na+(калия и натрия) из клетки и в клетку. После деполяризации через определенное время клетки переходят в состояние покоя, восстанавливая свою исходную полярность (изнутри минус, снаружи плюс), этот процесс называетсяреполяризацией.
Электрический импульс последовательно распространяется по отделам сердца, вызывая деполяризацию клеток миокарда. Во время деполяризации часть клетки оказывается изнутри заряженной положительно, а часть — отрицательно. Возникает разность потенциалов. Когда вся клетка деполяризована или реполяризована, разность потенциалов отсутствует. Стадиидеполяризации соответствует сокращениеклетки (миокарда), а стадииреполяризации — расслабление. На ЭКГ записывается суммарная разность потенциалов от всех клеток миокарда, или, как ее называют,электродвижущая сила сердца(ЭДС сердца).
Таким образом, ЭКГ — проекция вектора ЭДС на линию отведения.
164. Основные отведения электрокардиографии.
ЭКГ представляет собой запись изменения суммарного электрического потенциала, возникающего при возбуждении множества миокардиальных клеток.
Регистрация ЭКГ осуществляется с помощью электродов, накладываемых на различные участки тела. Система расположения электродов называется электрокардиографическими отведениями. При регистрации ЭКГ всегда используют 12 общепринятых отведений: 6-от конечностей и 6 грудных.
Первые 3 стандартных отведенияпредложены Эйнтховеном. Электроды накладывают следующим образом:
1 отведение левая рука (+) и правая рука (-),
2 отведение левая нога (+) и правая рука (-),
3 отведение левая нога (+) и левая рука (-).
Регистрируют также усиленные однополюсные отведения от конечностей:
аVR— от правой руки,
аVL— от левой руки,
аVF— левой ноги.
Усиленные отведения от конечностей находятся в определённом соотношении со стандартными.
Отведение аVLв норме имеет сходство c I отведением
aVR—cзеркально перевернутым II отведением
aVF— сходно с II и III отведениями.
6 грудных отведенийустанавливают на следующие точки
V1 — в 4 межреберье у правого края грудины,
V2 — в 4 межреберье у левого края грудины,
V3 — посередине между точкамиV2 иV4;
V4 — в 5 межреберье по левой срединно-ключичной линии;
V5 — на уровне отведенияV4 по левой передней аксиллярной линии;
V6 — на том же уровне по левой средней аксиллярной линии.
В некоторых случаях регистрируют дополнительные грудные отведения. К ним относятся V7,V8,V9, когда активный электрод располагается на уровнеV4-V6 соответственно по задней аксиллярной, лопаточной и паравертикулярной линиям.
165. Нормальная электрокардиограмма человека, ее генез, клиническое значение.
На нормальной ЭКГ имеется ряд зубцов и интервалов между ними. Выделяют зубец Р, зубцы Q,RиS, образующие комплексQRS, зубцыTиU, а также интервалыP-Q(P-R);S-T;Q-Т;Q-U;T-P.
Амплитуду зубцов измеряют в миливольтах. При этом 1 мВ соответствует отклонению от изолинии на 1 см.
Ширину зубцов и продолжительность интервалов измеряют в секундах по тому отведению, где эти параметры имеют наибольшую величину.
Генез ЭКГ:
1. Общее электрическое поле сердца образуется в результате сложения полей отдельных волокон сердечных мышц.
2. Каждое возбуждённое волокно представляет собой электрический диполь, обладающий элементарным дипольным вектором, характеризующийся определённой величиной и направлением.
3. Интегральный вектор в каждый момент процесса возбуждения представляет собой результирующую этих элементарных векторов.
4. Дипольный вектор направлен от (-) к (+), то есть от возбуждённого участка к невозбуждённому.
В каждый момент процесса возбуждения сердца отдельные векторы суммируются и образуют интегральный вектор. Возбуждение начинается в сино-атриальном узле, но оно на ЭКГ не отражается и поэтому записывается изолиния. Как только возбуждение переходит на предсердия, сразу же возникает разность потенциалов и на ЭКГ записывается восходящая часть зубца Р, отражающего возбуждение правого предсердия.
Возбуждение левого предсердия отражает нисходящая часть зубца Р.
В период формирования зубца Р возбуждение распространяется преимущественно сверху вниз. Это обозначает, что большая часть отдельных векторов направлена к верхушке сердца и интегральный вектор в этот период имеет ту же ориентацию.
Когда оба предсердия полностью охвачены возбуждением, и оно распространяется по атрио-вентрикулярному узлу, на ЭКГ записывается, изолиния (сегмент PQ).
Далее возбуждение распространяется по проводящей системе желудочков, а затем на миокард желудочков. Возбуждение желудочков начинается с деполяризации левой поверхности межжелудочковой перегородки. При этом возникает интегральный вектор, направленный к основанию, которое формирует зубец Q.
Далее, по мере распространения возбуждения на миокард правого и большую часть миокарда левого желудочка, вектор меняет направление на противоположное, то есть к верхушке и формирует зубец R.
Через стенку желудочков возбуждение распространяется от эндокарда к перикарду. В последнюю очередь возбуждается участок левого желудочка в области его основания, при этом интегральный вектор будет направлен вправо и кзади (то есть в сторону задней стенки желудочка) и формирует зубец S.
Когда желудочки полностью охвачены возбуждением и разность потенциалов между различными их отделами отсутствует, на ЭКГ записывается изолиния (сегмент ST).
Реполяризация желудочков отражается зубцом Т, который формируется вектором, направленным вниз и влево, то есть в сторону верхушки и левого желудочка.
Процесс реполяризации миокарда желудочков протекает значительно медленнее, чем деполяризация.
Скорость реполяризации в разных отделах различна: в области верхушек она наступает раньше, чем у основания, а в субэпикардиальных слоях раньше, чем в субэндокардиальных.
Таким образом, направление зубцов на ЭКГотражаеториентацию интегрального вектора.
Когда вектор направлен к верхушке сердца, на ЭКГ записываются (+), (направленные вверх) зубцы Р, R,Т.
Если вектор ориентирован к основанию, то записывается (-) зубцы QиS.

Треугольник Эйнтховена
Вольтаж зубцов в стандартном отведении имеет значение для определения положения электректрической оси сердца.
В норме электрическая ось сердца совпадает с анатомической и имеет направление
сзади-кпереди,
сверху-вниз,
справа-налево.
При этом наибольшую амплитуду зубцы имеют во IIотведении, так как оно отводит самую высокую разность потенциалов.
Высокий вольтаж зубцов в Iотведении свидетельствует о более горизонтальном расположении электрической оси сердца (горизонтальное сердце), а вIII— говорит о более вертикальном расположении электрической оси сердца (висячее сердце).
До настоящего времени нет общепринятой теории ЭКГ. Наиболее распространена дипольная теория. Она исходит из представлений о том, что границы между возбуждёнными и невозбуждёнными участками миокарда представляет собой линию, вдоль которой выстроен двойной слой электрических зарядов-диполей. На протяжении сердечного цикла за счёт распространения возбуждения по миокарду двойной электрический слой непрерывно перемещается, изменяет свою конфигурацию и в некоторые моменты может состоять из нескольких фрагментов.
Совокупность этих диполей можно представить в виде одного суммарного диполя, отражающего электродвижущую силу (ЭДС), сердца, величина и ориентация в пространстве которого непрерывно меняется. Потенциал точек, расположенных ближе к (+) полюсу диполя — (+), а потенциал точек, расположенных ближе к (-) полюсу — (-). Если точка одинаково удалена от обоих полюсов, то её потенциал=0.
Таким образом, ЭКГ — проекция вектора ЭДС на линию данного отведения.
Каждое отведение ЭКГ — это проекция электрической оси сердца (суммарного диполя) на соответствующую линию.
Стандартные отведения I,II,III— двухполюсные, то есть каждый из 2-х электродов — активный.
При I стандартном отведениирегистрируется разность потенциалов между правой и левой руками; то есть регистрируется суммарный диполь на фронтальную плоскость на линию «правая рука-левая рука».
Во II стандартном отведениирегистрируется разность потенциалов между правой и левой ногой; отражение идёт на фронтальную плоскость, но на линию расположенную под углом кI.
В III стандартном отведениипроизводится регистрация разности потенциалов между левой рукой и левой ногой; отражение идёт на фронтальную плоскость, на линию, которая соединяет левую руку и левую ногу.
Стандартные отведения позволяют определить расположение электрической оси (суммарного диполя) на фронтальной плоскости.
В нормальных условиях электрическая ось сердца расположена так, что она направлена справа - налево, сверху - внизи составляет по отношению к линии «правая рука - левая рука» (горизонтальная линия Эйнтховена) угол,=+20 -+70(такая позиция называетсянормограммой)RвоIIвыше, чем вIи особенно, чем вIII.
При горизонтальном положениисердца или гипертрофии левого желудочка электрическая ось сердца (суммарный диполь) изменяется — смещается влево. Угол с горизонтальной линией составляет меньше 20 градусов.
Признаком левограммыявляется значительное преобладание амплитуды зубцаRвIотведении над зубцамиRв остальных стандартных ответвлениях.
При гипертрофии правого желудочка, электрическая ось сердца (суммарный диполь) смещается вправо(правограмма) и угол становится больше 70 градусов. Признаком такого явления является преобладание по амплитуде зубцаRвIIIстандартном отведении над зубцамиRвIи воIIотведении.
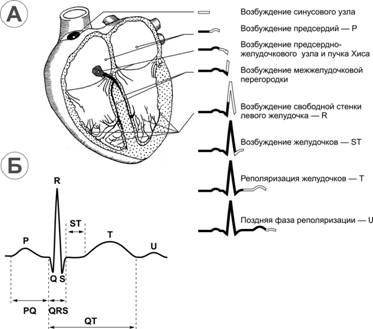
Генез ЭКГ
Нормальная ЭКГ человека
Зубец Р отражает возбуждение предсердий, положителен (направлен вверх) во всех отведениях, кромеaVR.
Может быть (-) в отведениях aVLилиaVFи отрицательным или двухфазным ( +-) в отведенияхV1 иV2.
Амплитуда не более 0,25 мВ, ширина — 0,1 с.
Первые 0,02-0,03 с отражают возбуждение правого предсердия, последние 0,02-0,03 с обусловлены только левопредсердным компонентом зубца.
Интервал P-Q
Отсчитывается от начала зубца Р до начала Q(при отсутствии — до началаR). Интервал отражает время, необходимое для деполяризации предсердий (зубец Р), проведение импульса через атриовентрикулярное соединение, пучок Гиса и его ветви.
Продолжительность интервала P-Qзависит от ЧСС (чем реже ритм, тем длиннее интервал). В норме не должен быть короче 0,12 с и не должен превышать 0,2 с.
Зубец Q
Представляет собой первый направленный вниз зубец желудочкового комплекса, предшествующий зубцу R, может отсутствовать. Отражает деполяризацию межжелудочковой перегородки.
В норме зубец Qможет быть глубоким, даже преобладающим в отведенииaVR.
Глубокий зубец Qв отведенияхIIIиaVFв сочетании с глубоким зубцомSвIиaVLможет наблюдаться при вертикальном положении оси сердца с поворотом вокруг продольной оси по часовой стрелке.
Выраженный Qвстречается вI,II,IIIотведениях при редко встречающемся положении сердца с поворотом верхушкой вперед.
Изредка в норме встречается зубец Q=50 % амплитуды зубцаRв отведенииaVL, а также зубецQSв отведенииV1.
В остальных отведениях в норме зубец Q не превышает по глубине 25 % амплитуды R
По ширине не более 0,03 с.
Зубец R
Отражает деполяризацию верхушки, передней, задней и боковой стенок желудочков сердца.
Высота от 0,5-2,5 мВ.
При расщеплении комплекса QRSи наличии нескольких зубцовRучитывается вершина последнего зубцаR.
Интервал Q-R
Время внутреннего отклонения (измеряется расстоянием от начала желудочкового комплекса (зубца QилиR) до проекции вершины зубцаRна изолинию.
Этот показатель характеризует время распространения возбуждения от эндокарда до эпикарда в месте расположения электрода, его используют для оценки последовательности возбуждения желудочков. В отведениях V1 иV2 (над правым желудочком) — 0,05 с.
Зубец S
Определяется как любой следующий за зубцом Rотрицательный зубец комплексаQRS. Этот зубец отражает процесс возбуждения основания желудочков сердца. Его амплитуда изменяется в широких пределах в зависимости от отведения, положения электрической оси сердца и других факторов. Максимальная глубина зубцаSв норме не более 2,5 мВ.
Комплекс QRS
Отражает деполяризацию желудочков. Ширину измеряют от начала зубца Qдо конца зубцаS. В норме не более 0,1 с. Максимальная амплитуда комплексаQRSв норме не более 2,6 мВ.
Сегмент S-T (R-T)
Отрезок от конца комплекса QRSдо начала зубца Т. Соответствует периоду угасания возбуждения желудочков и началу медленной реполяризации.
В норме — на изолинии; но в правых грудных отведениях может быть смещён вверх не более 0,2 мВ, в левых грудных отведениях - может быть смещён вниз не более 0,1 мВ.
Зубец Т
Отражает процесс быстрой реполяризации желудочков.
В норме — положителен во всех ответвлениях, кроме aVR, где он (-), иногда может быть (-) в отведенияхIIIиV1.
Амплитуда зубца Т в норме обычно составляет 1/8-2/3 амплитуды зубца R.
Ширина зубца Tколеблется от 0,1 до 0,25 с.
Интервал Q-T
Измеряется от начала зубца Q(R) до конца зубца Т. Соответствует электрической систоле желудочков. Его продолжительность зависит от ЧСС и других факторов.
Зубец U
Выявляется не всегда. В норме следует спустя 0,02-0,04 с после зубца Т. Отражает следовую реполяризацию волокон проводящей системы сердца. Амплитуда не более 0,25 мВ; продолжительность — 0,25 с.
Интервал Q-U
Измеряется от начала зубца Qдо концаU.
Интервал Т-Р
Отрезок ЭКГ от конца зубца Т до начала Р, интервал соответствует состоянию покоя миокарда.
166. Основные показатели деятельности сердца: частота и сила сердечных сокращений, систолический и минутный объемы крови в покое и при нагрузке.
Основной функцией сердца является нагнетание крови в систему сосудов. Насосная функция сердца характеризуется несколькими показателями. Одним из важнейших показателей работы сердца является минутный объем кровообращения (МОК) - количество крови, выбрасываемое желудочками сердца в минуту. МОК левого и правого желудочков одинаков. Синонимом понятия МОК является термин «сердечный выброс» (СВ). МОК - это интегральный показатель работы сердца, зависящий от величины систолического объема (СО) - количества крови (мл; л), выбрасываемого сердцем за одно сокращение, и ЧСС. Таким образом, МОК (л/мин) = СО (л) х ЧСС (уд/мин). В зависимости от характера деятельности человека в данный момент времени (особенности физической работы, поза, степень психоэмоционального напряжения и др.) доля вклада ЧСС и СО в изменения МОК различна. Ориентировочные величины ЧСС, СО и МОК в зависимости от положения тела, пола, физической подготовленности и уровня физической активности представлены в табл. 7.1.
Частота сердечных сокращений
ЧСС в покое. ЧСС - один из самых информативных показателей состояния не только сердечно-сосудистой системы, но и всего организма в целом. Начиная с рождения и до 20-30 лет ЧСС в покое снижается со 100-110 до 70 уд/мин у молодых нетренированных мужчин и до 75 уд/мин у женщин. В дальнейшем, с увеличением возраста, ЧСС незначительно возрастает: у 60-76-летних в покое по сравнению с молодыми на 5-8 уд/мин.
ЧСС при мышечной работе. Единственной возможностью повысить доставку кислорода к работающим мышцам является увеличение объема крови, поступающей к ним в единицу времени. Для этого должен возрасти МОК. Поскольку ЧСС прямо влияет на величину МОК, то повышение ЧСС при мышечной работе является обязательным механизмом, направленным на удовлетворение значительно возрастающих нужд метаболизма. Изменения ЧСС при работе показаны на рис. 7.6.
Если мощность циклической работы выразить через величину потребляемого кислорода (в процентах от величины максимального потребления кислорода - МПК), то ЧСС возрастает в линейной зависимости от мощности работы (потребления Ог, рис. 7.7). У женщин при условии равного с мужчинами потребления Ог ЧСС обычно на 10-12 уд/мин выше.
Наличие прямо пропорциональной зависимости между мощностью работы и величиной ЧСС делает частоту пульса важным информативным показателем в практической деятельности тренера и педагога. При многих видах мышечной деятельности ЧСС - точный и легкоопределяемый показатель интенсивности выполняемых физических нагрузок, физиологической стоимости работы, особенностей протекания периодов восстановления.
Для практических нужд необходимо знать величину максимальной ЧСС у лиц разного пола и возраста. С возрастом максимальные величины ЧСС как у мужчин, так и у женщин снижаются (рис. 7.8.). Точную величину ЧСС у каждого конкретного человека можно определить лишь опытным путем, регистрируя частоту пульса во время работы возрастающей мощности на велоэргометре. Практически для ориентировочного суждения о максимальной ЧСС человека (независимо от пола) используют формулу: ЧССмаКс = 220 - возраст (в годах).
Систолический объем сердца
Систолический (ударный) объем сердца - это количество крови, выбрасываемое каждым желудочком за одно сокращение. Наряду с ЧСС СО оказывает существенное влияние на величину МОК. У взрослых мужчин СО может меняться от 60-70 до 120-190 мл, а у женщин - от 40-50 до 90-150 мл (см. табл. 7.1).
СО - это разность между конечно-диастолическим и конечно-систолическим объемами. Следовательно, увеличение СО может происходить как посредством большего заполнения полостей желудочков в диастолу (увеличение конечно-диастолического объема), так и посредством увеличения силы сокращения и уменьшения количества крови, остающейся в желудочках в конце систолы (уменьшение конечно-систолического объема). Изменения СО при мышечной работе. В самом начале работы из-за относительной инертности механизмов, приводящих к увеличению кровоснабжения скелетных мышц, венозный возврат возрастает сравнительно медленно. В это время увеличение СО происходит в основном благодаря увеличению силы сокращения миокарда и уменьшению конечно-систолического объема. По мере продолжения циклической работы, выполняемой в вертикальном положении тела, благодаря значительному увеличению потока крови через работающие мышцы и активации мышечного насоса, возрастает венозный возврат к сердцу. Вследствие этого конечно-диастолический объем желудочков у нетренированных лиц со 120-130 мл в покое повышается до 160-170 мл, а у хорошо тренированных спортсменов даже до 200-220 мл. В это же время происходит увеличение силы сокращения сердечной мышцы. Это, в свою очередь, приводит к более полному опорожнению желудочков во время систолы. Конечно-систолический объем при очень тяжелой мышечной работе может уменьшиться у нетренированных до 40 мл, а у тренированных до 10-30 мл. То есть увеличение конечно-диастолического объема и уменьшение конечно-систолического приводят к значительному повышению СО (рис. 7.9).
В зависимости от мощности работы (потребления О2) происходят довольно характерные изменения СО. У нетренированных людей СО максимально увеличивается по сравнению с его уровне м в покое на 50-60%. У большинства людей при работе на велоэргометре СО достигает своего максимума при нагрузках с потреблением кислорода на уровне 40-50% от МПК (см. рис. 7.7). Иначе говоря, при увеличении интенсивности (мощности) циклической работы в механизме увеличения МОК в первую очередь используется более экономичный путь увеличения выброса крови сердцем за каждую систолу. Этот механизм исчерпывает свои резервы при ЧСС, равной 130-140 уд/мин.
У нетренированных людей максимальные величины СО уменьшаются с возрастом (см. рис. 7.8). У людей старше 50 лет, выполняющих работу с тем же уровнем потребления кислорода, что и 20-летние, СО на 15-25% меньше. Можно считать, что возрастное уменьшение СО является результатом снижения сократительной функции сердца и, по-видимому, уменьшения скорости расслабления сердечной мышцы.
Минутный объем кровообращения
Важным показателем состояния сердца является минутный объем кровотока, или минутный объем кровообращения (МОК). Нередко используют синоним понятия МОК - сердечный выброс (СВ). Величина МОК, являясь производной от СО и ЧСС (МОК = СО х ЧСС), зависит от многих факторов (см. табл. 7.1). Среди них ведущее значение имеют размеры сердца, состояние энергетического обмена в покое, положение тела в пространстве, уровень тренированности, величины физического или психоэмоционального напряжения, вид работы (статическая или динамическая), объем активных мышц.
В покое в положении лежа МОК у нетренированных и тренированных мужчин составляет 4,0-5,5 л/мин, а у женщин - 3,0-4,5 л/мин (см. табл. 7.1). В связи с тем, что МОК зависит от размера тела, при необходимости сравнения МОК у людей разного веса используют относительный показатель - сердечный индекс - отношение величины МОК (в л/мин) к площади поверхности тела (в м2). Площадь поверхности тела определяют по специальной номограмме, исходя из данных о весе и росте человека. У здорового человека в условиях основного обмена сердечный индекс обычно равен 2,5-3,5 л/мин/м2. В некоторых ситуациях (например, при низкой температуре окружающей среды) даже в условиях физического покоя возрастает энергетический обмен в организме. Это приводит к возрастанию ЧСС и, соответственно, МОК.
В положении стоя у всех людей МОК обычно на 25-30% меньше, чем лежа (см. табл. 7.1). Это связано с тем, что в вертикальном положении тела значительные объемы крови скапливаются в нижней половине туловища. Вследствие этого заметно уменьшается СО.
МОК и общий объем циркулирующей крови. Общий объем крови, находящейся в кровеносных сосудах, называется объемом циркулирующей крови (ОЦК). ОЦК - это важный параметр, определяющий давление, при котором происходит наполнение сердца кровью во время диастолы, а значит, и величину систолического объема. Величина ОЦК может претерпевать значительные изменения при переходе тела человека в вертикальное положение, при мышечных нагрузках, при воздействиях гормональных факторов, изменениях степени тренированности, окружающей температуры и т.д.
У взрослого человека около 84% всей крови находится в большом круге, 9% - в малом (легочном) круге и 7% - в сердце. Около 60-70% всей крови содержится в венозных сосудах.
Изменение МОК при мышечной работе. В условиях мышечной деятельности запросы мышц в кислороде возрастают пропорционально мощности выполняемой работы. При этом общее потребление организмом кислорода может возрастать в 10 и более раз. Вполне естественно, что это требует значительного увеличения МОК. Зависимость между величиной потребления кислорода (или мощностью работы) и МОК, вплоть до его предельных величин, носит линейный характер (см. рис. 7.7). Как уже отмечалось, МОК зависит от величины СО и ЧСС (МОК = СО х ЧСС). При мышечной работе увеличение МОК обусловлено возрастанием как СО, так и ЧСС. Конкретная величина МОК зависит от многих факторов. В частности, при одинаковой мощности работы в позе сидя или стоя МОК меньше, чем при работе в горизонтальном положении (рис. 7.10). При предельных аэробных нагрузках МОК у тренированных мужчин и женщин значительно выше, чем у нетренированных. Максимальные величины МОК у нетренированных мужчин и женщин уменьшаются с возрастом (см. рис. 7.8). При прочих равных условиях (пол, возраст, тренированность, положение исследуемого, окружающая температура и другие факторы) МОК зависит от объема активной мышечной массы и характера выполняемой работы. При динамической работе, в которой участвуют небольшие мышечные группы (например, работа одной или двумя руками), МОК меньше, чем при работе более'крупных мышц ног. При статической работе в отличие от динамической МОК почти не меняется. Это связано с тем, что кровообращение в мышцах практически прекращено. Приток крови к сердцу либо не меняется, либо даже может уменьшаться. Небольшие увеличения МОК, которые отмечают при изометрических сокращениях, связаны с заметным увеличением ЧСС при такого рода работе.
167. Тоны сердца, верхушечный толчок, их происхождение и характеристика. Фонокардиография, ее клиническое значение.
Тоны сердца –короткие отрывистые звуки, возникающие в здоровом сердце во время работы.
Тон – это звук, обусловленный периодическими колебаниями.
Простой (чистый) тон – это звук, обусловленный колебаниями одной частоты.
Сложный тон –совокупность простых тонов с кратным отношением частот.