

Руководство по иммунофармакологии / 013306
.pdf50 |
Глава 4 |
|
|
|
|
ний) и 5-окситриптамин (у человека) или |
нулах, находятся следующие: катепсины А, С и |
||||
гистамин (у кролика). 5-Окситриптамин не |
D; коллагеназа; эластаза; beta-глюкуронидаза; |
||||
синтезируется тромбоцитами, но, по-видимому, |
beta-N-ацетилглюкозаминидаза; |
beta- |
|||
захватывается ими и хранится в плотных |
глюкозидаза; Р-галактозидаза; а-галактозидаза; |
||||
гранулах. АТФ плотных гранул не входит в |
а-глюкози-даза; |
|
а-маннозидаза; |
а- |
|
метаболический пул АТФ тромбоцитов, и его |
арабинозидаза; |
|
beta-кси-лозидаза; |
||
обмен осуществляется крайне медленно по |
арилсульфатаза; beta-глицерофосфа-таза. |
|
|||
сравнению с обменом обычного пула АТФ. |
Тромбоциты способны изменять свою форму |
||||
а-Гранулы |
благодаря (в определенной мере) наличию |
||||
микрофиламентов |
и |
микротрубочек |
в |
||
Это гетерогенная группа гранул, содержащих |
цитоплазме. Микротрубочки располагаются по |
||||
периферии цитоплазмы и поддерживают форму |
|||||
фибриноген, фактор 4 (ФТ4) и его предшест- |
диска у интактных тромбоцитов. Мик- |
||||
венник, набор основных белков с разными |
рофиламенты содержат контрактильные белки и |
||||
формами активности (хемотаксической, усили- |
принимают участие в агрегации. Помимо |
||||
вающей рост, повышающей проницаемость), а |
различных форм актина и миозина, в тром- |
||||
также различные гликопротеины, в том числе |
боцитах содержатся регуляторные белки, на- |
||||
фибронектин и тромбоспондин. Исследование |
пример киназа легкой цепи, кальмодулин, |
||||
состава а-гранул показало, что часть их со- |
гельзолин и профилин. Основные структуры |
||||
держимого является результатом экзоцитоза из |
тромбоцита представлены на рис. 26. |
|
|||
окружающей среды. Однако фибриноген, |
|
|
|
|
|
входящий в состав этих гранул, по-видимому, |
Реакции тромбоцитов и их |
|
|||
отличается от плазменного и может синте- |
|
||||
зироваться мегакариоцитами, из которых про- |
определение |
|
|
|
|
исходят тромбоциты. |
Тромбоциты способны к следующим реакциям: |
||||
|
|
||||
Гранулы, содержащие кислую гидролазу |
1) изменение формы; 2) адгезия; 3) агрегация; 4) |
||||
Данные гранулы морфологически неотличимы |
выделение (или секреция) содержимого гранул; |
||||
5) выделение метаболитов ара-хидоната. В |
|||||
от а-гранул, но, вероятно, они не выделяются из |
зависимости от интенсивности и природы |
||||
тромбоцитов в результате секреции. Среди |
стимула общий ответ тромбоцитов может |
||||
ферментов, идентифицированных в этих гра- |
включать все перечисленные реакции или |
||||
|
|
только некоторые из них. Величина лю- |
|
||
Аморфное покрытие
Плотные гранулы
Мембрана
|
|
|
|
|
|
Микротрубочки |
|
|
|
|
|
|
|
|
|
|
Отложение |
|
|
|
|
||
|
|
|
|
|
|
|
|
Рис. 26. Внутриклеточные структуры |
Митохондрия |
Плотнотубулярная |
|||||
|
|
|
система |
|
|||
|
|
|
|
||||
тромбоцита.
гликогена
бого конкретного ответа также определяется типом стимула.
Изменение формы и адгезия
Изменение формы тромбоцита заключается в переходе из нормальной дисковидной формы в неправильную, с вдавлениями на поверхности и псевдоподиями. Реакция в виде изменения формы может наблюдаться и без выделения или агрегации и в отличие от других реакций не зависит от внетромбоцитарного кальция или изменений внутритромбоцитар-ного уровня циклического АМФ. Адгезия тромбоцитов к поверхности сопровождается образованием псевдоподий и изменениями формы, которые аналогичны индуцируемым стимулом в отсутствие какой-либо поверхности.
Агрегация
Агрегация, или прикрепление тромбоцитов друг к другу с образованием агрегата, в отличие от реакции изменения формы зависит от внутритромбоцитарного кальция и наличия фибриногена на поверхности тромбоцитов. Первая фаза агрегации (см. ниже) наблюдается, когда под влиянием стимула происходят конформационные изменения в паре поверхностных белков-гликопротеинов IIb/IIIa Конформационные изменения позволяют данным белкам связывать фибриноген, который является двухвалентным лигандом и, следовательно, может вызвать агрегацию тромбоцитов в результате образования перекрестных связей. Для второй фазы агрегации требуется секреция а-гранул, что зависит от образования в тромбоцитах тромбоксана А2. а-Гранулы содержат тромбоспондин, который при выделении связывается с фибриногеном первой фазы агрегации, стабилизирует агрегат и усиливает агрегацию.
Секреция
Секреция тромбоцитов обнаруживает немало сходства с другими секреторными процессами и не сводится к простому выделению из тромбоцита в результате его лизиса, а представляет собой экзоцитоз, при котором содержимое гранул селективно высвобождается из тромбоцита в окружающую среду. Действительно, выделение из гранул различных типов может происходить независимр, оно определяется
Тромбоциты 51
лишь природой стимула. Высвобождение плотных гранул иногда относят к секреции I, тогда как выброс из гранул, содержащих ферменты,- к секреции II. Секреция I связана с метаболизмом арахидоната и, вероятно, требует синтеза простагландиновых эндопереки-сей, так как она невозможна у тромбоцитов, обработанных ацетилсалициловой кислотой (см. главы 10 и 25). Различные стимулы, вызывающие реакции тромбоцитов, включают АДФ, адреналин, 5-окситриптамин, ФАТ, тромбоксан А2 и ПГ G2/H2, а также тромбин и коллаген. Из этого списка ясно, что тромбоциты содержат (или могут синтезировать при стимуляции) вещества, которые сами являются стимуляторами тромбоцитов. Таким образом, некоторые стимулы могут вызвать прямой ответ тромбоцитов, который может накладываться на вторичный (или непрямой) ответ при выделении веществ, а именно: метаболитов арахидоната и АДФ.
Измерение агрегации
Классическим методом контроля реакций тромбоцитов является определение изменений оптической плотности перемешиваемой суспензии тромбоцитов. На рис. 27 показана кривая, полученная с помощью агрегометра, измеряющего оптическую плотность.
Реакция на АДФ приведена для примера ситуации, когда первоначальное уменьшение пропускания света через суспензию связано с изменением формы тромбоцитов. Аналогичные реакции с изменением формы были получены при применении тромбина и коллагена. При стимуляции тромбоцитов адреналином подобный эффект отсутствует. Временное уменьшение оптической плотности сменяется ее увеличением, о чем свидетельствует плато. Это начальное увеличение определяется прямым агрегирующим действием АДФ на тромбоциты и называется первой фазой агрегации. Позже происходит дальнейшее увеличение оптической плотности (вторая фаза агрегации), которое связано с действием агрегирующих веществ, выделяемых из тромбоцитов при стимуляции (см. ниже). Таким образом, вторая фаза агрегации вызывается опосредованно и в случае АДФ может отменяться ацетилсалициловой кислотой, что указывает на участие метаболитов арахидоната.
При других стимулах кривые агрегации имеют иной вид. Коллаген, например, индуцирует только вторую фазу агрегации после

52 Глава 4
Рис. 27. Типичная кривая агрегометрии при стимуляции суспензии тромбоцитов АДФ.
Вторая фаза агрегации
первоначального изменения формы и не вызывает какого-либо прямого агрегационного ответа. АДФ, 5-окситриптамин, адреналин, тромбин и перекиси простагландинов вызывают как прямую, так и непрямую (т. е. вторую фазу) реакции агрегации.
Рецепторы
Тромбин
Тромбин является ферментом, катализирующим превращение фибриногена в фибрин; в то же время это один из наиболее мощных стимуляторов тромбоцитов. Активация тромбоцитов тромбином специфична, зависит от концентрации и рН среды и подавляется ингибиторами тромбина, т.е. имеет характеристики ферментативной реакции. Тромбин вызывает активацию тромбоцитов достаточно быстро (выявление менее чем через 10 с). Тромбоциты могут сенсибилизироваться к высоким дозам тромбина после их обработки подпороговыми концентрациями фермента. Исследования связывания не принесли успеха в отношении идентификации тромбинового рецептора. Примерно 500 мест связывания на тромбоците имеют высокий аффинитет к тромбину (константа диссоциации около 1 нМ); существует еще 50000 мест насыщаемого связывания на каждом тромбоците (константа диссоциации около 100 нМ). Ненасыщаемое
связывание составляет 25-75% общего связывания. Ни одно из мест связывания, как было четко показано, не является необходимым для активации тромбоцитов. Кроме того, некоторые аналоги тромбина активируют тромбоциты без насыщения. При равновесии большая часть тромбина находится в свободной форме, а связанный тромбин с течением времени становится недиссоциируемым от мест его связывания. Возможным объяснением этому служит предположение о том, что тромбин активирует тромбоциты, оказывая ферментативное действие на некий тромбоцитарный субстрат; при этом тромбин изменяется или подвергается процессингу.
Возможным кандидатом на мишень для действия тромбина является очень небольшой компонент поверхности тромбоцита, известный как гликопротеин V, который гидроли-зируется при низких концентрациях тромбина. Установлено, что для полной активации тромбоцита необходим гидролиз менее 2% гликопротеина V и что реакция тромбоцитов определяется не общим количеством гидролизируемого субстрата, а скоростью гидролиза. Однако против концепции о гликопротеине V как о мишени для тромбина свидетельствует тот факт, что другие протеолитические ферменты, гидролизирующие гликопротеин V, не вызывают активации тромбоцитов, хотя возможно, что эти ферменты индуцируют гидролиз определенного типа, не сопряженный с реакцией тромбоцитов.
|
|
|
|
Тромбоциты |
53 |
||
АДФ |
|
зу. 2', 5'-Дидезоксиаденозин подавляет аденилат- |
|||||
АДФ, высвобождающийся из |
поврежденных |
циклазу, причем он не только сам не вызывает |
|||||
агрегации тромбоцитов, но и не потенцирует |
|||||||
(при травме) клеток в сосудистое русло, от- |
агрегационный эффект АДФ. Аналоги АДФ- |
||||||
носится к наиболее важным физиологическим |
АДФаS и АДФbetaS-индуцируют агрегацию, но |
||||||
стимулам тромбоцитов. Рецепторы к АДФ на |
не подавляют аденилатциклазу. При исполь- |
||||||
тромбоцитах отличаются от пуриновых рецеп- |
зовании ряда конкурентных антагонистов АДФ |
||||||
торов других клеток прежде всего тем, что для |
было показано, что значение рА2 |
для АДФ- |
|||||
них АТФ является антагонистом, а не |
индуцированной агрегации хорошо коррелирует |
||||||
агонистом, как в случае других клеток. При |
с рА2 для подавления аденилатциклазы, которое |
||||||
использовании [beta-32Р]-2-азидо-АДФ выявлен |
вызвано АДФ. Таким образом, один и тот же |
||||||
единственный тип связывания с плотностью |
АДФ-рецептор, вероятно, опосредует агрегацию |
||||||
около 500 мест на каждом тромбоците. Кон- |
и подавление активности аденилатциклазы, но |
||||||
станта диссоциации для подавления связывания |
подавление |
аденилатциклазы |
не |
опосредует |
|||
АТФ имеет близкое значение с константой |
агрегацию. |
|
|
|
|
|
|
диссоциации при подавлении АТФ АДФ-инду- |
|
|
|
|
|
|
|
цированной агрегации. АДФ-рецептор тром- |
Адренорецепторы |
|
|
|
|||
боцитов обладает высокой степенью стерео- |
Агрегация тромбоцитов человека адреналином |
||||||
специфичности и другими структурными осо- |
|||||||
бенностями. Гуанин-, уридин- и цитозинди- |
опосредуется через а-адренорецепторы. Инте- |
||||||
фосфат неспособны вызвать агрегацию тром- |
ресно, что агрегации, вызванной адреналином, |
||||||
боцитов. Для полной активности в отношении |
не предшествует реакция изменения формы (см. |
||||||
рецептора необходимы рибоза АДФ и 5-ди- |
выше). На тромбоцитах млекопитающих |
||||||
фосфатная цепь АДФ. Как уже упоминалось |
выявлены и р-адренорецепторы, опосредующие |
||||||
выше, АТФ и АМФ являются антагонистами. |
подавление агрегации. У неприматов адреналин |
||||||
Замены в 8-м и 6-м положениях пуринового |
не агрегирует тромбоциты, но усиливает агре- |
||||||
кольца снижают активность, а некоторые за- |
гацию, вызванную другими стимулами. |
|
|||||
мены во 2-м положении ее повышают. Так, 2- |
а-Адренорецепторы на тромбоцитах отно- |
||||||
хлор-АДФ и 2-азидо-АДФ являются более |
сятся к а2-подтипу и селективно блокируются |
||||||
мощными стимуляторами агрегации тромбо- |
а2-антагонистом - йохимбином. На основании |
||||||
цитов, чем сам АДФ. |
|
определенного |
антагонистического |
действия |
|||
Аденозин подавляет агрегацию тромбоци- |
клонидина в отношении а-рецептора тромбо- |
||||||
тов, вызванную АДФ. Это подавление не яв- |
цитов предполагается, что данный рецептор |
||||||
ляется специфичным для АДФ-индуцируемой |
относится к а3-подтипу. Однако в настоящее |
||||||
активации тромбоцитов, и рецепторы, опосре- |
время установлено, что клонидин действует как |
||||||
дующие ингибиторный эффект аденозина, от- |
частичный агонист, поэтому фармакология а- |
||||||
личаются от пуриновых рецепторов, опосре- |
адренорецептора |
тромбоцитов |
находится |
в |
|||
дующих действие АДФ на тромбоциты. Аде- |
полном соответствии с характеристиками а2- |
||||||
нозин, действуя через собственный рецептор, |
подтипа. a1-Адренорецептор также вносит свой |
||||||
активирует аденилатциклазу, в результате чего |
вклад в процесс агрегации, так как а1 -агонист - |
||||||
уровень циклического АМФ в тромбоцитах |
метоксамин |
- |
стимулирует |
агрегацию |
|||
повышается, приводя к подавлению реакций |
тромбоцитов, полученных от некоторых до- |
||||||
тромбоцитов. Если активация аденозинового |
норов. Изучение связывания с мечеными ли- |
||||||
рецептора приводит к стимуляции аденилат- |
гандами подтвердило выводы |
относительно |
|||||
циклазы, то АДФ через свой рецептор вы- |
существования на тромбоцитах а2-, но не а1 - |
||||||
зывает ее подавление. В качестве ингибитора |
адренорецепторов. |
|
|
|
|||
аденилатциклазы АДФ почти |
эквипотентен |
|
|
|
|
|
|
самому себе в качестве агрегирующего агента. |
Рецепторы к эйкозаноидам |
|
|
|
|||
Отсюда возникает вопрос о взаимосвязи этих |
Простагландины В1 и D2, а также проста-циклин |
||||||
эффектов. Однако значительная часть данных |
|||||||
свидетельствует не в пользу гипотезы об опо- |
(ПГ12) подавляют агрегацию тромбоцитов, |
||||||
средовании агрегации тромбоцитов подавле- |
стимулируя |
аденилатциклазу |
и |
повышая |
|||
нием аденилатциклазы и снижением уровня |
внутритромбоцитарную концентрацию цАМФ. |
||||||
цАМФ тромбоцитов. Вазопрессин агрегирует |
При использовании радиоактивных |
агонистов |
|||||
тромбоциты, но не подавляет аденилатцикла- |
наблюдается более сложная картина |
|
|
||||
5-1134
54 |
|
Глава 4 |
|
|
|
|
|
|
|
|
|
взаимоотношений. Для ПГ12 |
на тромбоцитах |
этому используются для идентификации и |
|||||||||
выявлено два типа мест связывания: высоко- |
классификации рецепторов для ФАТ. В на- |
||||||||||
аффинные с константой диссоциации (KD) око- |
стоящее время предполагается существование |
||||||||||
ло 10 нМ и низкоаффинные с KD, примерно |
более чем одного типа рецепторов, причем |
||||||||||
равной 1 мкМ. Связывание ПГE1 |
также проис- |
рецепторы тромбоцитов обладают сходством с |
|||||||||
ходит в двух местах при KD 60 нМ и 2 мкМ, a |
рецепторами других тканей. При использо- |
||||||||||
ПГD2 |
обладает одним местом связывания при |
вании меченного тритием ФАТ-ацетоэфира для |
|||||||||
KD |
50 нМ. Связанный ПГЕ1 |
замещается |
изучения связывания с тромбоцитами человека |
||||||||
немеченым ПГЕХ или ПГІ2, но он резистентен к |
выявлено два типа мест связывания: с высоким |
||||||||||
замещению ПГD2. Аналогично этому связанный |
аффинитетом |
(KD |
около 0,3 нМ) и с более |
||||||||
ПГІ2 |
вытесняется немечеными ПГЕ, и ПГІ2, но |
низким аффинитетом (KD примерно 10 нМ). |
|||||||||
не ПГD2. Эти данные свидетельствуют о |
|
|
|
|
|
||||||
связывании ПГЕ1 |
и ПГІ2 с общим рецептором, |
Механизм передачи сигнала |
|
||||||||
который отличается от рецептора для ПГD2. |
|
||||||||||
Такой |
вывод |
соответствует |
результатам |
Кальций |
|
|
|
|
|||
исследований с использованием антагонистов, |
|
|
|
|
|||||||
таких как ди-4-флоретинфосфат, которые |
Кальций, несомненно, играет важнейшую роль |
||||||||||
препятствуют антиагрегационному |
эффекту |
||||||||||
ПГD2, но не ПГІ2 и ПГЕХ. |
|
и ПГІ2, по- |
в превращении лиганд-рецепторного взаимо- |
||||||||
В отличие от ПГЕ1 ПГD2 |
действия на мембране тромбоцита в клеточную |
||||||||||
давляющих активацию тромбоцитов, тромбо- |
реакцию. Кальциевые ионофоры, |
такие как |
|||||||||
ксан А2(ТА2) вызывает агрегацию тромбоцитов. |
А23187 и иономицин, могут вызывать все |
||||||||||
ТА2 |
|
индуцирует |
полный |
набор |
реакций |
реакции тромбоцитов, изменение формы, агре- |
|||||
тромбоцитов: изменение формы, агрегацию и |
гацию и секрецию. Однако роль кальция за- |
||||||||||
секрецию. Тромбоциты при метаболизме ара- |
висит, во-первых, от стимула и, во-вторых, от |
||||||||||
хидоната образуют inter alia эндоперекиси и |
типа вызываемой реакции. Например, стиму- |
||||||||||
ТА2. Эндоперекиси самостоятельно активируют |
ляция а-адренорецепторов вызывает агрегацию |
||||||||||
тромбоциты; так, ПГН2 индуцирует агрегацию в |
и секрецию посредством механизмов, не- |
||||||||||
присутствии ингибитора тромбоксан-синтетазы- |
зависимых от кальция. Тромбин, с другой |
||||||||||
дазоксибена. |
|
|
|
|
стороны, индуцирует реакции тромбоцитов, |
||||||
При изучении связывания меченого аналога |
которые частично зависят от внеклеточного |
||||||||||
ПГН2 |
выявлено по крайней мере три типа мест |
кальция и в большей степени-от выделения |
|||||||||
связывания. На один из них с плотностью около |
кальция из внутриклеточных депо, тогда как |
||||||||||
1700 мест на каждом тромбоците приходится |
реакция на АДФ в основном определяется |
||||||||||
более |
половины |
максимального |
связывания |
внеклеточным кальцием. |
|
|
|||||
лиганда в концентрации 70 нМ; по-видимому, |
Ценная информация была получена при |
||||||||||
он имеет наибольшую вероятность представлять |
измерении внутритромбоцитарной |
концентра- |
|||||||||
рецептор, что основывается на сравнении |
ции свободного кальция, [Ca2+]i, с помощью |
||||||||||
активности агониста и антагониста при |
индикатора квин-2. Изменение формы начи- |
||||||||||
вытеснении связанного лиганда и при действии |
нается при превышении [Ca2+]i |
порогового |
|||||||||
на агрегацию тромбоцитов. |
|
|
|
значения (около 300 нМ), а макс+имальный эф- |
|||||||
|
|
|
|
|
|
|
фект наблюдается при [Ca2+]i, |
равном 800 нМ. |
|||
Факторактивациитромбоцитов |
|
|
При таких уровнях кальция агрегация и секре- |
||||||||
Фактор активации тромбоцитов- 1-0-алкил-2- |
ция не регистрируются. Следует отметить, что |
||||||||||
некоторые стимуляторы тромбоцитов, такие как |
|||||||||||
ацетил-sn-глицерил-3-фосфорилхолин |
(ФАТ- |
АДФ и тромбин, могут вызвать изменение |
|||||||||
ацетоэфир, или ФАТ)-вызывает изменение |
формы при [Са2+]; |
ниже 300 нМ. Из этого можно |
|||||||||
формы, агрегацию и секрецию тромбоцитов; |
предположить |
существование |
второго |
||||||||
однако он способен, несмотря на свое название, |
передатчика. |
|
|
|
|
||||||
оказывать действие и на многие другие типы |
Первичная агрегация (или первая фаза аг- |
||||||||||
клеток (см. главу 16). Синтезированы |
регации) предполагает наличие на мембране |
||||||||||
антагонисты ФАТ, многие из которых обладают |
тромбоцитов |
фибриногеновых |
рецепторов в |
||||||||
селективностью; некоторые из них являются |
виде гликопротеинов IIb/IIIa. Для воз- |
||||||||||
конкурентными антагонистами и по- |
|
никновения |
первичной агрегации |
уровень |
|||||||
|
|
|
|
|
Тромбоциты |
55 |
|
[Ca2+]i должен повыситься примерно до 1 мкМ. |
повышается с помощью ионофоров от базаль- |
||||||
В то время как подобное повышение [Ca2+]i |
ного уровня (100 нМ) до 600 нМ (уровень |
||||||
может вызвать первичную агрегацию, как в |
слишком низкий для индукции секреции), а |
||||||
случае изменения формы, некоторые стимулы |
протеинкиназа С стимулируется ОАГ или ТФА, |
||||||
индуцируют первую фазу агрегации без |
то начинается секреция, количественные и |
||||||
увеличения [Са2+]i |
до 1 мкМ. Если [Са2+]; |
временные характеристики которой аналогичны |
|||||
повышается ионофором приблизительно до 600 |
наблюдаемым при секреции, индуцированной |
||||||
нМ, наблюдается изменение формы, но не |
физиологическим |
стимулом, |
например |
||||
агрегация. Добавление АДФ или тромбина в |
тромбином. Поэтому вполне вероятно, что |
||||||
этих условиях может вызвать агрегацию без |
стимуляторы тромбоцитов (тромбин, ФАТ, а |
||||||
дальнейшего увеличения [Ca2 + ]j, что опять- |
возможно, и АДФ) пробуждают тромбоциты к |
||||||
таки указывает на участие второго передатчика. |
секреции посредством механизма передачи, в |
||||||
Для секреции гранул тромбоцитов также |
котором принимают |
участие как |
повышение |
||||
требуется повышение [Са2+]i |
примерно до 1 |
[Са2+];, так и активация протеинкиназы С под |
|||||
мкМ; как и в случае изменения формы и |
действием ДАГ. Оба вторичных передатчика |
||||||
первичной агрегации, индукция секреции |
действуют синергично. На рис. 28 дана схема |
||||||
тромбином, ФАТ и другими стимулами воз- |
вторичных передатчиков, участвующих в со- |
||||||
можна при уровне [Са2+]i |
намного ниже 1 мкМ, |
пряжении процесса передачи сигнала на эф- |
|||||
что также свидетельствует в пользу участия |
фектор. |
|
|
|
|||
второго передатчика. |
|
|
Становится ясным, что для агрегации |
||||
|
|
|
|
тромбоцитов необходимо участие кальция и |
|||
Фосфоинозитиды |
|
|
|
ДАГ в качестве вторичных мессенджеров, од- |
|||
|
|
|
|
нако это, по-видимому, не дает полной картины |
|||
Как и у многих других клеток, взаимодействие |
процесса. Например, при использовании АДФ |
||||||
лиганд-рецептор на мембране тромбоцитов |
для индукции секреция не возникает в |
||||||
активирует фосфолипазу С, которая влияет на |
присутствии ацетилсалициловой кислоты или в |
||||||
фосфатидилинозитол, фосфатидилинозитол-4- |
отсутствие фибриногена. Это наблюдение |
||||||
фосфат и фосфатидилинозитол-4,5-дифосфат, в |
свидетельствует о том, что для проявления |
||||||
результате чего образуются |
диацилглицерол |
секреции, вызываемой АДФ, необходимы как |
|||||
(ДАГ), инозитол-1,4,5-трифосфат (ИФ3), ино- |
агрегация, так и обмен арахидоната через цик- |
||||||
зитол-1,4-дифосфат и инозитол-1-фосфат. ИФ3 |
лооксигеназный путь. Действительно, судя по |
||||||
действует как второй передатчик, выделяя |
имеющимся данным, вызванная АДФ первичная |
||||||
кальций из внутриклеточных депо и повышая |
агрегация необходима для начала образования |
||||||
[Са2+]i, а ДАГ активирует протеинкиназу С. |
ТА2 в тромбоцитах. ТА2 затем стимулирует |
||||||
Активация тромбоцитов тромбином, ФАТ и |
тромбоциты через собственный рецептор, что |
||||||
тромбоксаном А2 |
вызывает гидролиз фос- |
облегчает образование ДАГ в результате |
|||||
фоинозитидов с образованием ИФ3 и ДАГ. |
гидролиза фосфоинозитидов. ДАГ действует |
||||||
Данные относительно |
индукции гидролиза |
синергично с повышением [Ca2+]і, |
вызванным |
||||
фосфоинозитидов под действием АДФ весьма |
АДФ, и эти два сигнала индуцируют секрецию |
||||||
противоречивы. Возможно, АДФ вызывает |
(рис. 29). |
|
|
|
|||
гидролиз, но в меньшей степени, чем, напри- |
Важное значение проведенных исследований |
||||||
мер, тромбин. |
|
|
|
заключается в следующем: а) установлено |
|||
Как уже отмечалось, секреция тромбоцитов |
участие кальция и ДАГ как вторичных пе- |
||||||
в ответ на тромбин и ФАТ может происходить |
редатчиков; б) показана взаимосвязь различных |
||||||
при подпороговых уровнях [Ca2+]i ниже 1 мкМ, |
реакций тромбоцитов. И природа вторичных |
||||||
что предполагает участие второго передатчика. |
передатчиков, и характер взаимосвязи реакций |
||||||
Фармакологическая |
|
|
стимуляция |
определяются типом лиганд-рецеп-торного |
|||
протеинкиназы С 12-0-тетрадеканоил-форбол- |
взаимодействия, генерирующего реакцию. |
|
|||||
13-ацетатом (ТФА) или синтетическим олеоил- |
ЦиклическийАМФ |
|
|
|
|||
ацетилглицеролом (ОАГ) вызывает секрецию |
|
|
|
||||
тромбоцитов, но реакция начинается позднее и |
Как указывалось ранее, аденозин, ПГЕ2 и ПГІ2 |
||||||
протекает медленнее, чем при использовании |
|||||||
тромбина или других стимулов. Если [Ca2+]i |
вызывают в тромбоцитах повышение уровня |
||||||
|
|
|
|
цАМФ, что сопровождается подавлением pe- |
|
||

56 Глава 4
Рис. 28. Механизмы превращения сигнала в реакциях тромбоцитов.
Плюс-стимуляция; минус-подавление; ФЛС-фосфолипаза С; ФИФ2- фосфатидилинозитол ИФ3 - инозитол-1,4,5-трифосфат.
акций тромбоцитов. Предполагают, что цАМФ- |
рый не подавляет аденилатциклазу), в большее |
||||||
зависимые |
протеинкиназы |
тромбоцитов |
степени, чем агрегацию, индуцированнук АДФ |
||||
активируют |
кальциевые |
АТФазы, |
которые |
(подавляет |
аденилатциклазу). |
Возможно |
|
«качают» кальций либо из тромбоцитов, либо в |
определенный |
уровень внутритромбоцитарно- |
|||||
плотную тубулярную систему и другие |
го цАМФ модулирует реакции тромбоцитов, |
||||||
внутриклеточные места хранения кальция (см. |
однако его падение не служит вторичным пе- |
||||||
рис. 28). Кроме того, цАМФ-зависимые про- |
редающим сигналом, соединяющим рецептор- |
||||||
теинкиназы ограничивают вход кальция в ци- |
ную активацию мембран и реакцию, поскольку |
||||||
топлазму тромбоцитов или из внутритромбо- |
оно не является необходимым для индукции |
||||||
цитарных депо, или из внетромбоцитарной |
реакции. |
|
|
||||
жидкости. |
Внутритромбоцитарный |
цАМФ |
|
|
|
||
может служить модулятором реакций тромбо- |
Тромбоциты и воспаление |
|
|||||
цитов, которые вызываются кальцием (вто- |
|
||||||
ричным передатчиком). Возможно также, что |
Из предыдущих разделов главы понятно; что |
||||||
цАМФ как таковой является вторичным пе- |
|||||||
редатчиком для превращения активации мем- |
тромбоциты, как и тучные клетки или нейтро- |
||||||
бран в реакцию тромбоцитов; снижение внут- |
филы, служат источником ряда веществ, ко- |
||||||
ритромбоцитарного цАМФ |
необходимо перед |
торые могут опосредовать или регулировать |
|||||
повышением уровня кальция в цитозоле для |
воспалительные реакции. Более того, в местах |
||||||
активации реакций. Выше уже излагались фак- |
воспаления обнаруживаются вещества, акти- |
||||||
ты, свидетельствующие против этих положений. |
вирующие тромбоциты. Однако роль тромбо- |
||||||
Тем не менее ПГЕ1 и аденозин угнетают |
цитов в воспалении остается спорной; еще не |
||||||
активацию, вызванную вазопрессином (кото- |
накоплено достаточно данных, свидетельству- |
||||||
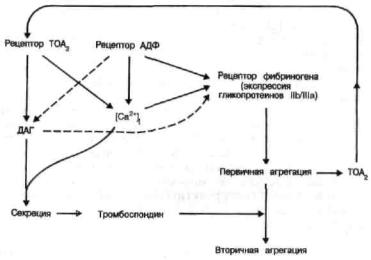
Тромбоциты 57
Рис. 29. Последовательность событий при опосредованных АДФ реакциях тромбоцитов [Hallam et alIn: Mechanisms of StimulusResponse Coupling in Platelets/ Eds. J. Westwick, M. F. Scully, D.E. Maclntyre, V. V. KakkarNew York: Plenum Press, 1985].
ющих о необходимости участия тромбоцитов в |
меняет реакции некоторых типов клеток, |
||||
воспалении. Следует, однако, повторить, что, |
участвующих в воспалении. ПГЕ2 вызывает |
||||
учитывая многокомпонентность воспаления, |
продолжительную гипералгезию, способству- |
||||
такие доказательства получить трудно. |
ющую усилению боли при воспалении. Таким |
||||
|
образом, накопление тромбоцитов в местах |
||||
Медиаторытромбоцитов |
воспаления и наличие в них медиаторов, об- |
||||
Показано, что при различных типах воспа- |
ладающих |
провоспалительными |
свойствами, |
||
могут служить косвенным доказательством их |
|||||
ления тромбоциты агрегируют и выделяют |
участия в процессах воспаления. |
|
|
||
содержимое своих гранул. Выделившиеся ги- |
|
|
|
|
|
стамин и 5-окситриптамин (в зависимости от |
Удалениетромбоцитов |
|
|
||
вида) способны увеличивать сосудистую про- |
Что же происходит с воспалительными реак- |
||||
ницаемость, а ферменты, такие как катепсины, |
|||||
эластаза и коллагеназа, могут повреждать вос- |
циями при удалении тромбоцитов? Острые |
||||
паленные ткани. Основные белки, также вы- |
воспалительные реакции у крыс в этих условиях |
||||
деляемые тромбоцитами, усиливают местную |
не изменяются. У тромбоцитопениче-ских крыс |
||||
сосудистую проницаемость. Показано, что ан- |
развиваются нормальный карраге-ниновый отек |
||||
тигениндуцированный бронхоспазм сопровож- |
лап и пассивная кожная анафилактическая |
||||
дается выделением в циркуляцию некоторых |
реакция. Интересно, что анти-тромбоцитарная |
||||
маркеров активации тромбоцитов, например |
сыворотка вызывает воспалительные реакции у |
||||
фактора 4 тромбоцитов. |
тромбоцитопенических крыс, причем картина |
||||
Активация тромбоцитов сопровождается |
воспаления не отличается от наблюдаемой у |
||||
увеличением активности фосфолипазы А2 и |
здоровых |
животных. |
|
Эффект |
|
арахидоната, образуемого под действием это- |
антитромбоцитарной |
сыворотки |
связан с |
||
го фермента, что приводит к повышению |
перекрестной реакцией с нетромбоцитар-ными |
||||
уровня циклооксигеназных и липоксигеназных |
антигенами. Приведенные данные показывают |
||||
продуктов (см. главу 10). Показано, что тром- |
важность |
определения |
специфичности |
||
боциты, стимулированные в местах воспале- |
антисыворотки до |
получения |
окончательных |
||
ния, образуют липоксигеназные вещества с |
результатов экспериментов с целевым ис- |
||||
хемотаксической активностью (ЛТВ4) и цикло- |
пользованием антисыворотки. В кожной пас- |
||||
оксигеназные продукты, способные модифи- |
сивной анафилактической реакции, вызванной |
||||
цировать воспалительные реакции (см. главы |
введением IgE-антител, не выявлено участия |
||||
6,7, 10, 14). ПГЕ2, например, усиливает воспа- |
тромбоцитов в отличие от реакции, индуци- |
||||
лительные эффекты других медиаторов и из- |
рованной |
преципитирующими |
|
антителами |
|
58 |
Глава 4 |
|
|
|
|
|
|
|
|
|
|
(IgG), |
которая |
сопровождается |
агрегацией |
местах реакции Артюса или в зонах поражения |
|||||||
тромбоцитов. Преципитирующие антитела в |
при сывороточной болезни. В этой связи важно |
||||||||||
отличие от IgE способны фиксировать ком- |
отметить специфические характеристики кро- |
||||||||||
племент, что, вероятно, и вызывает активацию |
ликов, у которых тромбоциты содержат только |
||||||||||
при данном типе реакции пассивной кожной |
гистамин, в отличие от тромбоцитов человека и |
||||||||||
анафилаксии. |
|
сыворотка |
вызывает |
крыс, содержащих 5-окситриптамин. Кроме |
|||||||
Антитромбоцитарная |
того, у кроликов отсутствуют IgE-анти-тела, |
||||||||||
спазм бронхов, что, вероятно, связано с тром- |
поэтому активация тромбоцитов в реакциях |
||||||||||
боцитами, поскольку у животных с низким |
повышенной чувствительности осуществляется |
||||||||||
уровнем тромбоцитов бронхоспазм не возни- |
преципитирующими антителами (IgG), которые, |
||||||||||
кает. У кроликов анафилактический шок ха- |
как уже указывалось, фиксируют комплемент. |
||||||||||
рактеризуется наличием тромбоцитарных эм- |
Тромбоцитопения |
сопровождает |
приступы |
||||||||
болов в легких, а гепарин оказывает некоторое |
астмы у человека; при этом обнаружены |
||||||||||
защитное действие. У морских свинок ФАТ |
циркулирующие |
комплексы |
|
агрегатов |
|||||||
обусловливает |
гиперреактивность |
бронхов, |
тромбоцитов. |
|
|
|
|
|
|||
которая аналогична наблюдаемой при брон- |
Эндотоксин, липополисахарид стенок грам- |
||||||||||
хиальной астме у человека, причем данный |
отрицательных бактерий, вызывает агрегацию и |
||||||||||
эффект ФАТ зависит от тромбоцитов. Кроме |
секрецию у тромбоцитов. Кроме того, после |
||||||||||
того, у морских свинок ФАТ вызывает брон- |
инъекции эндотоксина тромбоциты в виде аг- |
||||||||||
хоспазм, зависимый от тромбоцитов. В на- |
регатов накапливаются в микрососудах. Вве- |
||||||||||
стоящее время установлено существование ре- |
дение эндотоксина вызывает шок, а препараты, |
||||||||||
цепторов к IgE на тромбоцитах, и, хотя они |
подавляющие реакции тромбоцитов, в неко- |
||||||||||
отличаются от аналогичных рецепторов тучных |
торой степени защищают от шока. Введение |
||||||||||
клеток (см. главу 2), их присутствие предпо- |
второй дозы эндотоксина после примирующей |
||||||||||
лагает наличие механизма активации тромбо- |
или сенсибилизирующей инъекции приводит к |
||||||||||
цитов при взаимодействии аллерген - IgE. Ан- |
развитию состояния, напоминающего дис- |
||||||||||
тиастматический препарат хромогликат по- |
семинированную |
внутрисосудистую |
коагуля- |
||||||||
давляет в тромбоцитах IgE-зависимое генери- |
цию. Тромбоциты, таким образом, участвуют в |
||||||||||
рование свободных радикалов, которые явля- |
воспалительных и других (сосудистых) реак- |
||||||||||
ются |
важным медиатором гиперреактивности |
циях на эндотоксин. |
|
|
|
|
|||||
бронхов. ФАТ индуцирует гиперреактивность |
Тромбоциты, по-видимому, способны само- |
||||||||||
бронхов, что отличает его от других стиму- |
стоятельно вызывать гибель бактерий и ви- |
||||||||||
ляторов тромбоцитов; поэтому возможно, что |
русов, а они в свою очередь могут индуцировать |
||||||||||
данный эффект ФАТ не связан с тромбоцитами. |
агрегацию и секрецию у тромбоцитов. |
||||||||||
Тромбоцитопения уменьшает эозинофиль-ную |
Тромбоциты могут выделять |
бактерицидное |
|||||||||
инфильтрацию легких, которая обусловлена |
вещество 0-лизин, а стимуляция тромбоцитов |
||||||||||
введением аллергена |
сенсибилизированным |
комплексом антиген - IgE приводит к высво- |
|||||||||
животным, что свидетельствует об участии |
бождению из тромбоцитов достаточно боль- |
||||||||||
эозинофилов в формировании гиперреак- |
шого количества |
цитотоксических |
свободных |
||||||||
тивности дыхательных путей. Однако до сих |
радикалов, вызывающих гибель паразитов. |
||||||||||
пор неизвестна степень участия тромбоцитов в |
Протеины, выделяющиеся из активированных |
||||||||||
развитии бронхиальной астмы у человека. |
таким образом тромбоцитов, могут повреждать |
||||||||||
|
|
|
|
|
и разрушать бактерии. Микроорганизмы могут |
||||||
Тромбоцитыприразличныхтипах |
|
захватываться тромбоцитами, но значение этого |
|||||||||
воспаления |
|
|
|
процесса |
не |
определено. |
Тромбоциты, |
||||
По имеющимся данным, тромбоциты (у кро- |
поглотившие микроорганизмы, по-видимому, |
||||||||||
более |
быстро |
захватываются |
|
клетками |
|||||||
ликов) не только способствуют некоторым |
ретикулоэндотелиальной |
системы, |
что, |
||||||||
острым воспалительным реакциям, но и при- |
безусловно, ускоряет очищение циркуляции от |
||||||||||
нимают участие в реакциях замедленного типа, |
вирусов и других микроорганизмов. |
|
|
||||||||
таких как реакция Артюса и сывороточная |
Клеточно-опосредованные иммунные реак- |
||||||||||
болезнь. Свидетельства их участия представ- |
ции (например, реакция «трансплантат против |
||||||||||
лены в основном морфологическими данными, |
хозяина») осуществляются с участием тромбо- |
||||||||||
показывающими |
секрецию тромбоцитов в |
цитов. Тромбоцитарные тромбы являются ха- |
|||||||||
|
|
|
|
|
рактерным признаком в |
зоне |
отторжения |
||||
Тромбоциты 59
трансплантата. Значение этого факта опять-таки не определено; возможно, оно заключается в реакции на повреждение тканей, которое вызвано клетками и химическими медиаторами отторжения.
В заключение следует отметить, что тромбоциты во многих отношениях сходны с другими клетками, участвующими в воспалительных и иммунных реакциях. Они содержат (или могут синтезировать) мощные медиаторы воспаления и вовлекаются в такие реакции соответствующими стимулами. Тем не менее роль тромбоцитов в воспалительных реакциях до конца не выяснена, а представленные доказательства их участия в подобных реакциях в значительной мере приблизительны.