

системные механизмы
.pdfбета-1 в 1,11±0,04 раза и уменьшение амлитуды дельта ритма301 в 1,18±0,05 раза. Частоты генерации бета-2 статистически значимо возросли в 1,01±0,0014 раза, а альфа ритма в 1,01±0,0028 раза. Произошла реорганизация ЭКоГ, состоявшая в статистически значимом уменьшении индексов длительности бета-2 и тета ритмов в 1,28±0,03 и 1,06±0,03 раза с оответственно.
В правом полушарии наблюдалось (табл. 8.10) статистически значимое увеличение амплитуд бета-2 ритма, бета-1 и альфа ритмов в
1,22±0,04, 1,26±0,05 1,20±0,05 раз соответственно.
Статистически значимых изменений частот ритмов ЭкоГ после совместного применения БП и ПФ не наблюдалось. Реорганизация ЭКоГ состояла в статистически значимом уменьшении длительности бета-2 и тета ритмов в 1,38±0,03 и 1,13±0,02 раз соответственно и увеличении длительности дельта ритма в 1,15±0,04 раза раз соответственно.
Изменения показателей ФМПА ЭКоГ после исчезновения судорожной активности, в условиях введения ПФв. До введения БП показатели ФМПА (табл. 8.11, рис. 8.12) амплитуды ритмов ЭКоГ находились в пределах (2,36±0,04%)
– (5,05±0,19%), т.е были латерализованы в левое полушарие. После введения БП и ПФ и исчезновения судорожной активности положительность показателя ФМПА амплитуды бета-2 ритма ЭКоГ уменьшилась до –0,60±0,03, а бета –1, альфа, тета и дельта ритмов выражались отрицательными величинами, т.е. инвертировали. До введения БП показатели ФМПА частоты ритмов ЭКоГ выражались отрицательными величинами в пределах (-0,03± -0,00008) -(-0,31±-0,001).
После введения БП и ПФв и прекращения судорожной активности показатели ФМПА частоты бета-2, альфа и тета ритмов ритма инвертировал и выражались положительными величинами, т.е латерализовали в левое полушарие. Показатели ФМПА частоты бета-1 и дельта ритмов увеличилм отрицательность.
Показатели ФМПА индексов длительности бета-2, альфа и дельта ритмов ЭКоГ до введения БП и ПФв выражались положительными значениями 1,22±0,03%, 1,18±0,04% и 2,83±0,09% соответственно, а бета-1 и тета ритмов – отрицательными значениями -1,97±0,04%, -3,77±0,07%

соответственно. После исчезновения судорожной активности302 произошло увеличение положительности показателей ФМПА индексов длительности бета-2 и альфа ритмов и уменьшение отрицпательности бета-1 и тета ритмов. Показатель ФМПА индекса длительности дельта ритма инвертировал из положительного в отрицательный.
Tаблица 8.9. Показатели ЭКоГ и статистически значимые
коэффициенты соотношения левого полушария до и после исчезновения судорожной активности индуцированной внутрибрюшинным введением БП, в условиях применения ПФв.
Показател |
|
До |
После |
Статистически |
и ЭКоГ |
|
введения |
введения БП |
значимые |
|
|
БП |
|
коэффициенты |
|
|
|
|
соотношения |
Бета-2 |
А |
47,44±0,46 |
55,99±1,71* |
1,18±0,04 |
|
Ч |
27,81±0,03 |
27,98±0,03* |
1,01±0,00 |
|
И |
18,52±0,28 |
14,51±0,25* |
-1,28±0,03 |
Бета-1 |
А |
85,38±1,60 |
95,13±2,98* |
1,11±0,04 |
|
Ч |
16,07±0,02 |
16,02±0,03 |
|
|
И |
13,62±0,20 |
13,78±0,29 |
|
Альфа |
А |
124,70±3,35 |
134,58±3,81 |
|
|
Ч |
9,74±0,02 |
9,84±0,02* |
1,01±0,00 |
|
И |
17,52±0,45 |
18,94±0,49 |
|
|
А |
171,79±3,39 |
163,24±4,01 |
|
Тета |
Ч |
5,52±0,01 |
5,57±0,02 |
|
|
И |
33,54±0,49 |
31,54±0,58* |
-1,06±0,03 |
Дельта |
А |
250,17±7,15 |
211,88±5,81* |
-1,18±0,05 |
|
Ч |
2,78±0,01 |
2,78±0,02 |
|
|
И |
37,11±0,90 |
38,14±1,00 |
|
Примечание: * P<0,05

Tаблица 8.10.303 Показатели ЭКоГ и статистически значимые
коэффициенты соотношения правого полушария до и после исчезновения судорожной активности индуцированной внутрибрюшинным введением БП, в условиях применения ПФв.
Показатели |
До |
После |
Статистически |
||
ЭКоГ |
|
введения |
введения БП |
значимые |
|
|
|
|
БП |
|
коэффициенты |
|
|
|
|
|
соотношения |
Бета-2 |
|
А |
45,24±0,56 |
55,32±1,92* |
1,22±0,04 |
|
|
Ч |
27,89±0,02 |
27,81±0,03 |
|
|
|
И |
18,07±0,25 |
13,11±0,18* |
-1,38±0,03 |
Бета-1 |
|
А |
78,19±1,48 |
98,13±3,68* |
1,26±0,05 |
|
|
Ч |
16,14±0,02 |
16,14±0,02 |
|
|
|
И |
14,17±0,22 |
13,94±0,18 |
|
Альфа |
|
А |
114,46±3,14 |
137,33±4,55* |
-1,20±0,05 |
|
|
Ч |
9,75±0,02 |
9,78±0,02 |
|
|
|
И |
17,11±0,38 |
16,89±0,51 |
|
|
|
А |
162,96±3,79 |
176,88±4,60 |
|
Тета |
|
Ч |
5,55±0,01 |
5,52±0,01 |
|
|
|
И |
36,17±0,45 |
32,09±0,48* |
-1,13±0,02 |
Дельта |
|
А |
226,10±5,25 |
224,72±5,85 |
|
|
|
Ч |
2,79±0,01 |
2,81±0,01 |
|
|
|
И |
35,07±0,75 |
40,32±0,97* |
1,15±0,04 |
Примечание: * P<0,05
Таблица 8.11. Изменения показателей ФМПА ЭКоГ до введения БП и
после исчезновения судорожной активности, индуцированной внутрибрюшинным введением БП, в условиях предварительного введения ПФв.
Показатели |
До введения |
После |
|
Эког |
БП и ПФв |
исчезнове- |
|
|
|
|
ния |
|
|
|
судорожных |
|
|
|
потенциалов |
Бета-2 |
А |
2,36±0,04 |
0,60±0,03* |
|
Ч |
-0,13±0,00 |
0,29±0,00* |
|
И |
1,22±0,03 |
5,06±0,11* |
|
|
|
|

Продолжение таблицы 8.11304
Бета-1 |
А |
4,40±0,12 |
-1,55±0,08* |
|
Ч |
-0,21±0,00 |
-0,35±0,00* |
|
И |
-1,97±0,04 |
-0,56±0,01* |
Альфа |
А |
4,28±0,16 |
-1,01±0,04* |
|
Ч |
-0,03±0,00 |
0,31±0,00* |
|
И |
1,18±0,04 |
5,70±0,23* |
|
А |
2,64±0,08 |
-4,01±0,14* |
Тета |
Ч |
-0,31±0,00 |
0,38±0,00* |
|
И |
-3,77±0,07 |
-0,87±0,02* |
Дельта |
А |
5,05±0,19 |
-2,94±0,11* |
|
Ч |
-0,24±0,00 |
-0,47±0,00* |
|
И |
2,83±0,09 |
-2,77±0,10* |
|
|
|
|
Примечание: * P<0,05
8.2.2.3. Эволюция ФМПА и лобно-затылочного коэффициента судорожной активности вызванной
внутрибрюшинным введением БП и ПФв.
В условиях совместного применения БП и ПФв ФМПА судорожной активности биологической модели
6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
% |
β1 α |
||||||||||||||
|
|
|
|
|
|||||||||||||
4 |
β2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
θ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0
-2
-4
-6
до введения пенициллина
δ
β2
 α β1
α β1
δ
θ
после исчезновения судорог
показатели
Рисунок 8.12. Показатели ФМПА амплитуды ритмов ЭКоГ до введения БП и ПФ и после исчезновения судорожной активности
многоочаговой корковой эпилепсии была выражена меньшими значениями, как положительности -12,36±0,15%,
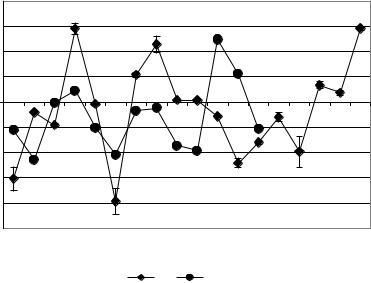
так и отрицательности (-11,40±0,08%), чем при305 изолированном применении БП (рис. 8.13). В течение эволюции биологической модели при применении ПФв отмечено три цикла по 80 минут.
20 |
|
|
|
|
|
|
|
|
% |
|
|
|
|
15 |
|
|
|
|
|
|
10 |
|
|
|
|
|
|
5 |
|
|
|
|
|
|
0 |
|
|
|
|
|
|
-5 |
|
|
|
|
|
|
-10 |
|
|
|
|
|
|
-15 |
|
|
|
|
|
|
-20 |
|
|
|
|
|
|
-25 |
|
|
|
|
|
|
0 |
20 |
40 |
60 |
80 |
100 120 140 160 180 200 220 240 260 280 300 320 340 |
|
|
|
|
|
|
Время (мин) |
|
|
|
|
|
|
БП |
БП+ПФ |
Рисунок 8.13. Динамика ФМПА судорожной активности вызванной внутрибрюшинным введением БП.
Обозначения: БП – применение БП, БП+ПФв – применение БП и ПФв.
В условиях применения ПФв величины лобнозатылочного коэффициента (рис. 8.14) находилась в течение
200 минут в пределах 4,16±0,54% - (-7,58±1,84%). В период исчезновения судорожной активности к 220 и 240 минутам наблюдения отрицательность величины лобно-затылочного коэффициента возросла (-15,95±3,86%) и (-33,09±10,03%).
8.3. Взаимоотношения амплитуд ритмов ЭКоГ до формирования и после прекращения судорожной
активности.
До введения БП в левом полушарии (рис. 8.15, А) определялось 8 положительных связей-отношений, а в правом (рис. 8.15, Б) – 10 положительных и 2 отрицательные. Таким образом в левом полушарии всего было 8 связейотношений, а в правом – 12. После окончания эволюции судорожной активности индуцированной внутрибрюшинным введением БП, в левом полушарии (рис. 8.15, Г) определялось

7 положительных и 1 отрицательная связь-отношение, а в306 правом (рис. 8.15, Д) – 11 положительных и 3 отрицательные связи-
60 |
|
% |
|
|
|
|
|
|
|
|
|
|
|
40 |
|
|
|
|
|
|
20 |
|
|
|
|
|
|
0 |
|
|
|
|
|
|
-20 |
|
|
|
|
|
|
-40 |
|
|
|
|
|
|
-60 |
|
|
|
|
|
|
0 |
20 |
40 |
60 |
80 |
100 120 140 160 180 200 220 240 260 280 300 320 340 |
|
|
|
|
|
|
Время (мин) |
|
|
|
|
|
|
БП |
БП+ПФ |
Рисунок 8.14. Динамика лобно-затылочного коэффициента судорожной активности вызванной внутрибрюшинным введением БП.
Обозначения: БП – применение БП, БП+ПФ – применение БП и ПФ.
отношения. Таким образом в левом полушарии всего было 8 связей-отношений, а в правом – 14.
До формирования судорожной активности и введения ПФв в левом полушарии (рис. 8.16, А) определялось 10 положительных и две отрицательные связи-отношения, а в правом (рис. 8.16, Б) – 8 положительных и 8 отрицательных.
Таким образом в левом полушарии всего было 12 связей-отношений, а в правом – 16. После окончания эволюции судорожной активности индуцированной внутрибрюшинным введением БП в условиях предварительного введения ПФ, в левом полушарии (рис. 8.16, Г) определялось 6 положительных и 4 отрицательные связи-отношения, а в правом (рис. 8.16, Д) – 8 положительных. В левом полушарии всего было 10 связейотношений, а в правом – 8.
Таким образом после окончания эволюции судорожной активности, индуцированной внутрибрюшинным введением БП в левом полушарии количество связей-отношений не изменилось, а в правом полушарии определялось на две

связи-отношения больше, чем до начала эволюции307 судорожной активности. После окончания эволюции судорожной активности индуцированной внутрибрюшинным введением БП в условиях дополнительного введения ПФв, количество связей-отношений в левом полушарии уменьшилось на две, а в правом на 8 связей-отношений.
\
А |
Б |
|
|
В |
|
Г |
Рисунок |
8.15. |
Матетическая |
модель |
отражающая |
взаимоотношения между амплитудами ритмов ЭКоГ крыс до и после введения БП .
Обозначения: А – до введения левое поушарие Б – до ведения правое полушарие, В - после прекращения судорожой активности левое поушарие, Г - после прекращения судорожой активности правое полушарие.
1 – бета –2 ритм, 2 – бета 1 ритм, 3 – альфа ритм, 4 – тета ритм, 5 – дельта ритм.
Следует отметить, что в правом полушарии после окончания эволюции судорожной активности индуцированной внутрибрюшинным введением БП в

условиях дополнительного введения ПФ, не определялись308 отрицательные связи-отношения.
В целом проведенные опыты свидетельствуют о наличии у ПФв противосудорожного эффекта, выявленного, как на модели первично-генерализованной эпилепсии сформированной бемегридом, так и на модели корковой эпилепсии сформированной внутрибрюшинным введением БП.
А |
Б |
В |
Г |
Рисунок 8.16. Математическая модель отражающая взаимоотношения между амплитудами ритмов ЭКоГ левого полушария крыс до и после введения ПФв и БП .
Обозначения: А – до введения левое поушарие Б – до ведения правое полушарие, В - после прекращения судорожой активности левое поушарие, Г - после прекращения судорожой активности правое полушарие.
1 – бета –2 ритм, 2 – бета 1 ритм, 3 – альфа ритм, 4 – тета ритм, 5 – дельта ритм.
Проведенные исследования показали, что ПФв, эномеланин, экстракт красной свеклы и экстракт белого винограда обладают
противосудорожным действием при испытании на309 биологической модели первично-генерализованной эпилепсии, сформированной применением коразола и бемегрида. Это может свидетельствовать о том, что противосудорожное действие флавоноидов полифенольной природы, является характерным признаком биологически активных препаратов этой группы.
Высказано предположение, что главный механизм иктально-межиктальной циркуляции в течение эпилептического приступа - чередующееся накопление в течение иктальной фазы и клиренса в течение межиктальной фазы эндогенного аденозина (Eldridge F.L., Paydarfar D., Scott S.C., Dowell R.T. 1989). С другой стороны, показано, что применение аденозина потенциирует противосудорожное действие ресвератрола и предположено, что противоэпилептический эффект ресвератрола реализуется использованием аденозинэргического механизма. (Gupta Y.K,.Chaudhary G., Srivastava A.K. 2002).
По-видимому, полученный в наших опытах противосудорожный эффект на первично-генерализованной биологической модели эпилепсии, сформированной пирименением бемегрида и коразола, и пенициллиновой модели эпилепсии можно объяснить, хотя-бы отчасти, использованием аденозинэргического механизма.
Однако выявленный противосудорожный эффект при применении антиоксидантов, в частности токоферола
(Ayyildiz M., Yildirim M., Agar E. 2006; Крижановский Г.Н.,
Тупеев И.Р., Никушкин Е.В. 1987), позволяют предположить что антиоксидантное действие ПФв также вносит свой вклад в его противосудорожное действие. По-видимому, имеется несколько, возможно независимых, биологических механизмов действия ПФв.
Внутрибрюшинное введение БП в дозе 300 000 МЕ на 100 г. массы тела крыс вызывало формирование многоочаговой корковой эпилепсии с выраженными периодами интериктальной и иктальной судорожной активности.
Динамика амплитуд характеризовалась возрастанием величин судорожных потенциалов к 80 минуте наблюдения после введения БП. В этот период наблюдения максимальные величины амплитуд регистрировались в левом полушарии а
минимальные в затылочной коре. Затем с 100 минуты до 260310 минуты наблюдался относительно стабильный период генерирования амплитуд судорожных потенциалов. К 340 минуте наблюдения амплитуды судорожных потенциалов стремились к 0.
Изменение частоты генерации судорожной активности носила волнообразный характер. Максимальные значения частот генерации судорожных потенциалов, регистрировались на 180 минуте после введения БПа. Наибольшее значение частот генерации судороржных потенциалов достигала в затылочной коре, а наименьшее – во фронтальной коре.
Эволюция частоты и амплитуды судорожной активности в эпилептическом комплексе описывались уравнением вида y=a+bx+cx2lnx+dx0,5lnx+ex/lnx.
При моделировании с помощью данного уравнения эволюции эпилептического комплекса вне периода наблюдения – 360-540 минут было получено теоретическое возрастание амплитуд и и частот генерации судорожных потенциалов. Полученные результаты позволили предположить, что на 2-3 день после внутрибрюшинного введения БП у крыс может регистрироваться ЭпА.
При структурном анализе полициклических мультиграфов описывающих связи-отношения между показателями амплитуд судорожной активности зарегистрированной в различных регионах коры головного мозга выявилось, что наибольшее количество связейотношений наблюдалось к четвертому часу эволюции эпилептического комплекса. Количество статистически значимых коэффициентов корреляции в этот период наблюдалось наименьшее. В начале и в конце существования эпилептической активности количество связей-отношений определялось наименьшим, а количество статистически значимых коэффициентов корреляции наибольшим.
Подобные отношения между коэффициентами множественной регрессии и коэффициентами корреляции выявлялись и при расчетах показателей амплитуд ритмов ЭКоГ. Учитывая, что выявленная закономерность выявляется как в условиях фоновой ЭКоГ так и судорожной активности,