

системные механизмы
.pdfдлительности альфа и тета ритма в левом полушарии –231 сниженными.
Во фрагментах синхронизации применение ПФв в условиях АП, по сравнению с аналогичными фрагментами ЭКоГ, зарегистрированными в условиях удаления полушарий, но без применения ПФв (табл. 6.11, рис. 6.17), выявлено увеличению амплитиуд бета-2, бета-1, альфа и тета ритма в обеих полушариях и дельта ритма в правом полушарии. Индексы длительности бета- 1 и дельта ритмов определялись увеличенными, а альфа и тета ритмов – уменьшенными, но только в правом полушарии.
6.4.2. Изменения индексов ФМПА ЭКоГ при применении ПФв в условиях препарата с удаленными полушариями мозжечка.
Как во фрагментах десинхронизации, так и во фрагментах синхронизации ФМПА амплитуд ритмов ЭКоГ (табл. 6.12, рис. 6.18) выражалась отрицательными величинами. Отрицательность ФМПА амплитуд ритмов во фрагментах десинхронизации находилась в пределах (-5,60±0,11%) – (- 8,88±0,16%), а во фрагментах синхронизации в пределах – (- 22,69±0,72%) – (31,95±1,90%), т.е. была более выражена.
После введения ПФв во фрагментах десинхронизации ФМПА амплитуды бета-2 ритма статистически значимо не изменилась, а бета-1, альфа и тета ритмов статистически значимо увеличилась, а дельта ритма – уменьшилась. После введения ПФв во фрагментах синхронизации ФМПА амплитуд всех ритмов статистически значимо уменьшились.
Препарат «изолированного мозга» является достаточно корректной моделью для изучения механизмов действия фармакологических агентов (Лабори А., 1970; Крыжановский Г.Н и др., 1980). Характерным признаком ЭКоГ препарата изолированного мозга является увеличение амплитуды и индекса длительности альфа ритма в левом полушарии и латерализация показателей ЭКоГ в левое полушарие.
Особенности электрогенеза препарата изолированного мозга кошки было изучено ранее (Руссев В.В., Макулькин Р.Ф., 1959;
Серков Ф.Н., и др., 1960; 1963; Серков П.М. та iн., 1964).
Усиление латерализации ритмов ЭКоГ в левое полушарие на препарате изолированного мозга может свидетельствовать об
асимметрии собственных синхронизирующих механизмов232 левого и правого полушарий, а именно, большей активностью механизмов синхронизации левого полушария. По-видимому, в условиях интактного мозга различия в активности синхронизирующих механизмов нивелируются восходящими влияниями ретикулярной формации.
К 5 минуте после введения ПФв в ЭКоГ левого полушария отмечалось только увеличение показателя индекса длительности альфа ритма, а в правом увеличение амплитуд бета-2, бета-1 и альфа ритма, частоты и индекса длительности бета-2 и бета-1 ритмов.
350
300
250
200
150
100
50
0
500
400
300
200
100
0
А
мкВ
δ
θ
α
β1
β2
1 2 3 4 |
1 2 3 4 |
1 2 3 4 |
1 2 3 4 |
1 2 3 4 |
|||||
|
|
|
Б |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
мкВ |
|
|
|
|
|
|
|
δ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
θ
α
β1
β2
1 2 3 4 |
1 2 3 4 |
1 2 3 4 |
1 2 3 4 |
1 2 3 4 |
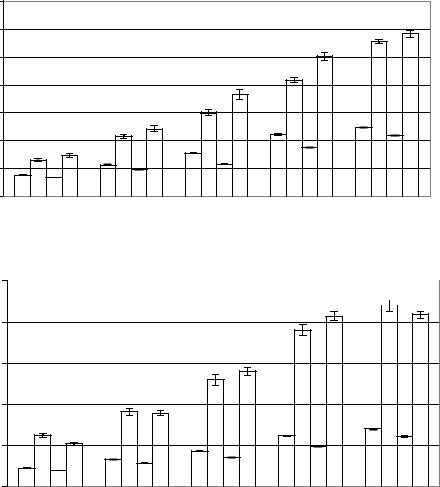
Рис. 6.17. Диаграммы средних амплитуд ритмов 2-, 1-, -, -, и-ЭКоГ, отводимой от левого (А) и правого (Б) полушарий до (1, 2) и после внутрибрюшинного введения ПФр (3, 4), в пределах

сегментов десинхронизации (1, 3) и синхронизации (2, 4), в233 условиях аспирации полушарий мозжечка.
К 10 минуте после введения ПФв в ЭКоГ левого полушария изменения отсутствовали, а в ЭКоГ правого полушария отмечалось увеличение показателей бета-2 ритмаамплитуды, частоты и индекса длительности.
К 15 минуте после введения ПФв в левом полушарии произошло уменьшение амплитуд всех диапазонов ЭКоГ, частот бета-2, бета-1 и дельта ритма и индексов длительности бета-1 и тета римов, а в правом полушарии наблюдалось только увеличение индекса длительности бета-2 ритма.
Таблица 6.12. Показатели ФМПА до и после введения ПФв в условиях
препарата изолированного переднего мозга при использовании процедуры сегментации
Показатели |
До введения ПФв |
После введения ПФв |
|||
ЭКоГ |
|
Десинхрони- |
Синхронии- |
Десинхрони- |
Синхронии- |
|
|
зация |
зация |
Зация |
зация |
Бета-2 |
А |
13,96±0,26 |
19,99±1,10 |
-2,23±0,02 |
-2,92±0,10 |
|
Ч |
3,32±0,03 |
6,44±0,14 |
-4,38±0,02 |
-2,78±0,02 |
|
И |
18,11±0,49 |
31,22±2,14 |
-6,80±0,08 |
4,67±0,14 |
Бета-1 |
А |
25,33±0,49 |
31,47±1,99 |
8,54±0,07 |
13,36±0,48 |
|
Ч |
1,76±0,01 |
5,70±0,08 |
-1,16±0,00 |
1,56±0,01 |
|
И |
10,81±0,22 |
31,62±1,62 |
-3,43±0,03 |
9,01±0,26 |
Альфа |
А |
23,07±0,43 |
28,90±2,14 |
15,12±0,13 |
13,05±0,65 |
|
Ч |
4,25±0,03 |
11,49±0,40 |
1,10±0,00 |
4,02±0,05 |
|
И |
15,81±0,36 |
28,56±2,32 |
4,63±0,05 |
10,43±0,40 |
Тета |
А |
12,29±0,20 |
0,05±0,00 |
19,66±0,13 |
17,02±0,57 |
|
|
|
|
|
|
|
Ч |
1,58±0,01 |
1,05±0,02 |
1,95±0,00 |
2,36±0,03 |
|
И |
5,32±0,10 |
2,16±0,13 |
8,22±0,07 |
5,02±0,17 |
Дельта |
А |
6,85±0,12 |
0,17±0,00 |
16,43±0,12 |
18,49±0,30 |
|
Ч |
-0,28±0,00 |
-4,61±0,10 |
-0,75±0,00 |
1,92±0,03 |
|
И |
-12,50±0,23 |
-5,27±0,17 |
-10,13±0,09 |
-3,95±0,07 |
|
|
|
|
|
|
С 20 по 40 минуты после введения ПФв в ЭКоГ левого полушария отмечалось уменьшение амплитуд, частот и индексов длительности бета-2 и бета-1 ритмов ЭКоГ.
Изменения ЭКоГ с 20 по 50 минуты наблюдения в правом полушарии были менеее выражены: к к 40 и 50 минутам

наблюдения отмечалось уменьшения индекса длительности234 бета-1 ритма, к 35 минуте отмечалось увеличение частоты, а к 50 амплитуды дельта ритма.
КА после применения ПФв в левом полушарии уменьшилась к 5, 15, 20, 25, 30, 35 и 40 минутам после применения ПФв, а в правом полушарии КА, напротив, увеличивался к 10, 15, 25 минутам после применения ПФв.
В целом можно отметить, что в левом полушарии после введения ПФв отмечено уменьшение уровня активации, а в правом – возрастание.
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
-5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
α θ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
θ δ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
δ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
-10 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β2 β1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β2 |
β1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
α |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
-15 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
-20 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
δ |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
α |
|
|
|
|
|
|
|
|
|
|||
-25 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
δ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
θ |
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
-30 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β1 |
|
α |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
% |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
θ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
-35 |
|
|
|
|
|
|
|
|
|
|
|
β2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
-40 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
|
2 |
|
|
|
|
|
|
|
|
|
|
|
|
3 |
|
|
|
|
|
|
|
|
|
4 |
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||
Рис. 6.18. Диаграммы коэффициентов функциональной межполушарной асимметрии амплитуд ритмов 2-, 1-, -, -, и ЭКоГ, отводимой до (1 и 2) и после внутрибрюшинного введения ПФв (3, 4), в пределах сегментов десинхронизации (1, 3) и синхронизации (2, 4) в условиях АП.
Таким образом, наиболее существенные изменения ЭКоГ препарата изолированного мозга после введения ПФв были
выявлены в отношении бета-2 и бета-1 ритмов. Следует235 подчеркнуть, что реакция на введения ПФв вначале регистрировалась в правом полушарии - к 5 и 10 минутам после введения, а в левом полушарии изменения ЭКоГ отмечены только после 10 минуты. Также следует отметить различный характер реагирования ЭКоГ на введение ПФв левого и правого полушарий: в левом полушарии уменьшение показателей ЭКоГ бета ритма, а в правом увеличение. Вполне понятно, что Ка в левом полушарии определялся уменьшенным по сравнению с исходным уровнем, а в правом увеличенным.
При разрушении латерокаудальной части покрышки среднего мозга содержание норадреналина в диенцефалоне и теленцефалоне уменьшается на 75%. Это свидетельствует о том, что значительная часть норадренергических терминалей в диенцефалоне и теленцефалоне происходят от клеточных тел, лежащих в мосту и продолговатом мозге (Буданцев А.Ю., 1976). Показано, что дофаминэргические терминали сосредочены в основном в определенных, ограниченных областях переднего мозга зонах(Буданцев А.Ю., 1976).
Активация дофаминовой системы сопровождается появлением в ЭЭГ высокочастотной бета-активности, которая снижается при приеме нейролептиков (Панюшкина С.В. с
соавт. 1994).
Выявленные после введения ПФв на препарате изолированного мозга изменения в диапазоне бета активности, и относительная интактность остальных диапазонов ЭКоГ, могут свидетельствовать, с одной стороны о тропности ПФв к дофамину, а с другой, о сохранности именно дофаминовых нейрохимических механизмов на препарате изолированного переднего мозга.
В левом полушарии бета активность после введения ПФв подавлялась, а в правом усиливалась. Полученный факт может свидетельствовать о нейрохимической межполушарной асимметрии во влиянии ПФв на дофаминэргичсекие нейрохимические механизмы, которые обусловлены, учитывая выявленную межполушарную асимметрию собственных синхронизирующих механизмов левого и правого полушарий, межполушарной асимметрией дофаминэргических систем левого и правого полушарий.
Между амплитудами ритмов ЭКоГ на препарате236 изолированного мозга определялось значительно большее, чем у интактных крыс количество взаимных ориентированных влияний – 4,33 (13/3) и 14,67 (44/3) в левом и правом полушариях соответственно у интактных животных, и 20 и 14 на препарате изолированного мозга. Следует отметить, существенное возрастание количества связей в левом полушарии.
После применения ПФв количество статистически значимых связей между амплитудами ритмов ЭКоГ левого полушария уменьшилось с 20 до 13, а правого полушария возросло с 14 до 16.
В условиях интактного мозга возрастание КА сопровождается уменьшением межамплитудных связей. В условиях препарата изолированного мозга, напротив, увеличение КА в правом полушарии сопровождалось увеличением количества связей, а уменьшение величин КА в левом полушарии, сопровождалось уменьшением межамплитудных взаимоотношений.
Изменения КА при применении ПФв в условиях препарата изолированного мозга были обусловлены изменениями бета активности, в то время как, в условиях интактного мозга в левом полушарии после применения ПФв изменения бета-2 индекса не определялись, а индекс времени альфа ритма определялся уменьшенным к 5, 20, 25 и 30 минутам после введения. В правом полушарии интактного мозга уменьшения альфа индекса отмечалось к 5, 10 и 15 минутам после введения, а уменьшения индекса длительности бета-2 ритма (совместно с уменьшение индекса длительности альфа римтма) к 15 минуте наблюдения.
Полученные различия могут быть обусловлены различными нейрофизиологическими механизмами формирования КА в условиях интактного мозга и препарата изолированного мозга. В условиях интактного мозга увеличение КА обеспечивается уменьшением индекса длительности альфа ритма, в то время как в условиях препарата изолированного мозга - влиянием на активность бета ритма. Снижение альфа индекса свидетельствует об усилении неспецифической церебральной активации и обусловлено активацией ретикулярной системы среднего мозга (Мельник
Э.В., 1998). Изменения альфа-индекса отражают уровень237 восходящей активации (Лукьянова Н.Ф., Малкин В.Б., 1977).
По-видимому, в условиях интактного мозга действие ПФв реализуется преимущественно через норадренергические механизмы ретикулярной формации, а в условиях препарата изолированного мозга – через дофаминэргические нейрохимические механизмы.
На препарате изолированного мозга остается сохранной ТЛ-продуцирующая система переднего гипоталамуса. Поэтому представляло интерес сопоставить ЭКоГ-эффекты после внутрибрюшинного применения ПФв в условиях препарата изолированного мозга и центрального применения ТЛ в левый и правый боковые желудочки мозга.
При корреляции показателей ЭКоГ на препарате изолированного мозга после применения ПФв и левостороннего центрального введения ТЛ статистически значимыми отрицательными определялись в левом полушарии корреляции между показателями амплитуд бета-2 и альфа ритмов, а в правом полушарии статистически значимо, положительно коррелировали изменения амплитуд тета ритмов.
При корреляции показателей ЭКоГ после применения ПФв и правостороннего центрального введения ТЛ статистически значимыми отрицательными определялись в левом полушарии коэффициенты корреляции между показателями бета- 1 ритма: амлитуды, частоты и индекса длительности, индекса длительности альфа ритма и частоты тета ритмов. В правом полушарии статистически значимыми отрицательными определялись изменения показателей индексов длительности тета ритма.
В правом полушарии статистически значимо отрицательно коррелировали изменения амплитуд альфа индекса и частоты тета ритмов ЭКоГ.
Также как, было выявлено в условиях интактного мозга, большинство статистически значимых коэффициентов корреляций показателей ЭКоГ при применении ПФв и ТЛ определялись в левом полушарии.
Следует отметить, что как в условиях интактного мозга, так и в условиях изолированного мозга при левостороннем применении ТЛ статистически значимо отрицательно
коррелировали изменения амплитуд бета-2 ритма, а при238 правостороннем применении ТЛ в левом полушарии статистически значимо отрицательно коррелировали изменения амплитуд бета-1 ритма.
Квадраты расстояний Маханолобиса между разностными матрицами показателей ЭКоГ при центральном левостороннем применении ТЛ и введении ПФв в условиях изолированного мозга составляли в левом полушарии 2,04 при Р=0,11, а в правом – 3,13 при Р=0,021. При центральном правостороннем применении ТЛ и введении ПФв в условиях изолированного мозга в левом полушарии квадраты расстояний Маханолобиса составляли в левых полушариях 2,43 при Р=0,066, а в правых
3,81 при Р=0,011.
Таким образом, подобие в изменении показателей ЭКоГ при применении ТЛ и ПФв в условиях изолированного мозга выявлялось только в левых полушариях, при центральном левостороннем применении ТЛ.
Следует отметить, что в условиях интактного мозга подобие в изменении показателей ЭКоГ при применении ТЛ и ПФв также было выявлено при левостороннем центральном применении ТЛ, но в левом и правом полушариях.
На препарате изолированного мозга ритмы ЭКоГ по амплитуде и частоте латерализованы в левое полушарие. Латерализация по индексам длительности ритмов ЭКоГ подобна таковой у интактных крыс – индексы длительности бета – 2, бета – 1, альфа и тета ритмы латерализованы в левом полушарии, а индекс дельта ритма - в правом, но выражается большими значениями.
После внутрибрюшинного введения ПФв в условиях препарата изолированного мозга положительность показателей ФМПА амплитуд ритмов статистически значимо уменьшилась, а показатель ФМПА амплитуды бета-2 ритма инвертировал и выражался отрицательной величиной.
После применения ПФв статистически значимо уменьшилась положительность показателей ФМПА частоты бета-2 и альфа ритмов ЭКоГ препарата изолированного мозга, а показатели ФМПА частоты бета-1, тета и дельта ритмов статистически значимо не изменились.
Положительность показателей ФМПА индексов длительности бета-2, бета-1 и альфа ритмов ЭКоГ препарата
изолированного мозга после применения ПФв статистически239 значимо уменьшилась, а тета и дельта ритмов определялась статистически значимо не измененной.
Таким образом, применение ПФв способствует уменьшению левосторонней латерализации ритмов ЭКоГ.
На препарате с удаленными полушариями мозжечка в левом полушарии после введения ПФв длительность фрагментов десинхронизации увеличилась на 9,00%, а длительность фрагментов синхронизации, напротив, уменьшилась на 6,53%. Длительность фрагментов ЭКоГ, графоэлементы которых представляли различные формы пароксизмальных проявлений уменьшилась на 2,47%. В правом полушарии после введения ПФв длительность фрагментов десинхронизации незначительно уменьшилась на 1,25%, а длительность фрагментов синхронизации, напротив, незначительно увеличилась на 0,92%. Длительность фрагментов ЭКоГ, графоэлементы которых представляли различные формы пароксизмальных проявлений незначительно увеличилась на 0,34%.
Таким образом, под влиянием ПФв временные изменения графоэлементов ЭКоГ выявлялись в левом, но не правом, полушарии. Ранее была высказана гипотеза (Лобасюк Б.А., 2005), что нейрофизиологическое действие ПФв может реализовываться через стимуляцию норадренергических и тиролиберинэргических нейрохимических механизмов, преимущественно левого полушария. Полученные результаты, в какой то мере, подтверждают эту гипотезу.
Реорганизация ЭКоГ при переходе от десинхронизации к синхронизации после примененимя ПФв в условиях удаления полушарий мозжечка выражалась, в частности, увеличением амплитуд ритмов, и это увеличение было более выражено, чем в аналогичной ситуации без применения ПФв. Во фрагментах десинхронизации снижение амплитуд ритмов ЭКоГ после примененимя ПФв в условиях удаления полушарий мозжечка было болем существенным, чем в аналогичной ситуации без применения ПФв.
Во фрагментах десинхронизации после примененимя ПФв по сравнению с аналогичными фрагментами до введения ПФв амплитуда всех ритмов в левом полушарии уменьшалась, а в правом амплітуда бета-2, бета-1 и альфа ритмов ЭКоГ
статистически значимо не изменилась, тета ритма240 уменьшилась, а дельта ритма – возросла. Во фрагментах синхронизации после примененимя ПФв по сравнению с аналогичными фрагментами до введения ПФв амплитуда всех ритмов как в левом, так и правом полушарии уввеличилась. При применении ПФв в условиях интактного мозга амплитуды ритмов ЭКоГ как во фрагментах десинхронизации, так и синхронизации уменьшалась.
Таким образом, действие ПФв в условиях удаления полушарий мозжечка, в периоды десинхронизации в левом, но не правом, полушарии реализуется также, как и в интактном мозге – подавлением амплитуд, а в периоды синхронизации результат действия ПФв противоположен, тому который наблюдается в интактном мозге и реализуется увеличением амплитуд ритмов ЭКоГ.
Можно высказать предположение, что уменьшение амплитуд в периоды десинхронизации реализуется путем активации ретикулярной формации, а более выраженное уменьшение амплитуд слева обусловлено более развитыми связями коры левого полушария, чем правого, со стволовыми структурами головного мозга.
Увеличение амплитуд ритмов ЭКоГ во фрагментах синхронизхации при применении ПФв в условиях удаления полушарий мозжечка, возможно, обусловлено реципрокными влияниями из левого полушария.
В целом, можно высказать предположение, что в условиях интактного мозга нейрофизиологическое действие ПФв реализуется с участием полушарий мозжечка.