

2. Строение мицелия
Мицелий – система ветвящихся трубок, гиф с апикальным ростом и боковым ветвлением. Мицелий может быть клеточный и неклеточный.
Неклеточный мицелий лишён перегородок. В процессе роста мицелия, деление ядер не сопровождается образованием клеточных перегородок – септ, что ведёт к развитию большой массы цитоплазмы, содержащей много ядер. Такой мицелий представляет собой одну гигантскую многоядерную клетку (ценоцит). С одной стороны, это хорошо, так как все части организма объединены в единое целое; с другой – плохо, даже небольшое повреждение может вызвать гибель значительной части таллома (пока пораженное место не будет отделено перегородкой). Ценоцит характерен для хитридиомикот, оомикот и зигомикот.
У некоторых хитридиевых наблюдается дифференциация ценоцита: от центральной части отходят нитевидные выросты, называемые ризомицелием. Часто эти выросты не содержат ядер и не могут выполнять много функций, но они более эффективно оккупируют субстрат, например, паразит водорослей Polyphagus eugienae с помощью ризомицелия может одновременно питаться несколькими клетками эвглен.
Возникновение ризомицелия создало большие возможности для морфологической эволюции грибов. У примитивных хитридиомицетов бесполый цикл происходит следующим образом: зооспора, попавшая на поверхность клетки растения, переливает содержимое в клетку хозяина, где превращается в амебоид. Ядра разросшегося амебоида делятся митотически; после чего он целиком превращается в спорангий и прекращает существование после освобождения зооспор. Подобный тип таллома называется холокарпическим, а спорангий — моноцентрическим. Развитие ризомицелия привело к миграции ядер в отдельные его участки (собирательные клетки), которые приобрели способность превращаться в зооспорангии. Таким образом, на одном талломе возникало несколько зооспорангиев, и таллом не погибал после формирования зооспор, а продолжал вегетировать. Такой эукарпический таллом является зачатком истинного мицелиального строения.
|
|
|
Рис. 1. Типы септ. А. Простая, Б. Долипоровая. 1 – пора, 2 – долипора, 3 – парентосома |
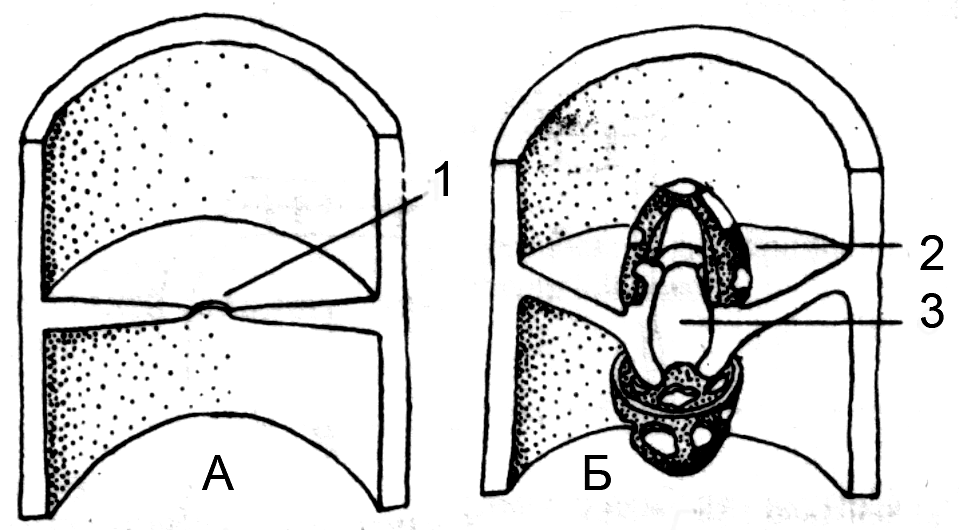
У большинства грибов развита простая септа в поперечной перегородке которой одна центральная пора, при этом септа утоньшается по направлению к поре — (рис. 1). У базидиальных грибов долипоровая септа, которая утолщается по направлению к поре. У некоторых грибов долипоровая септа несёт поровый колпачок – парентосому. Строение поры – один из важнейших признаков в систематике грибов.
Мицелиально-дрожжевой диморфизм
Многие виды грибов в разных условиях могут находиться в мицелиальном или дрожжевом состоянии, такое явление называется мицелиально-дрожжевой диморфизм. При переходе к дрожжевому росту происходит перестройка морфологии, физиологии и метаболизма грибов; изменяется физиология энергетических процессов (дыхание заменяет брожение); образ жизни (паразитический или сапротрофный); химический состав клеточной стенки (глюканы заменяются на маннаны) и цитоплазматической мембраны (содержание входящих в нее углеводов падает, а белка и липидов – увеличивается).
Причины диморфизма:
1. Физиология питания. Мицелиальные формы дейтеромицетов из рода Verticillium живут, как сапротрофы в почве, но могут заражать корни высших растений, проникать в сосуды ксилемы и существовать там в дрожжеподобной форме, вызывая увядание (вилт) пораженного растения. Другой дейтеромицет Candida albicans в дрожжевой форме паразитирует в позвоночных животных и человеке, вызывая поражения внутренних органов (глубокие микозы), а в мицелиальной форме живет в почве и питается растительными остатками.
2. Состояние ядер в клетке. Базидиальные грибы из порядка головневые образуют в зараженных тканях растений хорошо развитый дикариотический мицелий. Такой мицелий не может расти на искусственных питательных средах, он – облигатный паразит. Но одноядерные споры этих грибов размножаются почкованием, образуют дрожжеподобные колонии на искусственной среде и не способны заражать растения.
3. Состав среды. Некоторые зигомицеты из рода Mucor в аэробных условиях существуют в мицелиальной форме, а в анаэробных — в дрожжевой. Mycotypha africana (Zygomycetes) в питательной среде с 20% глюкозы при 37°С растет исключительно в форме почкующихся клеток («круглых дрожжей»), а при обычном содержании сахара (например, 2%) или более низких температурах (например, 27°С) – в виде гиф, быстро переходящих к спороношению.
Плектенхима.
При плотном переплетении гиф у грибов образуется ложная ткань, плектенхима (от греч. plektys — сплетенный, свитый). Плектенхима отличается (по происхождению и строению) от настоящей ткани, которая возникает в результате деления клеток и в поперечном, и в продольном направлениях. У грибов гифы мицелия делятся, как правило, с образованием только поперечных перегородок. Поэтому плектенхима непрочная, рыхлая, но именно из нее сложены плодовые тела, стромы и склероции грибов.