

3.3.2 Зависимость вероятности электронных переходов отcis[Ca]
При разработке
электронно-конформационной модели было
сделано предположение о зависимости
вероятности электронных переходов от
концентрации Са2+
в cis-части (cis[Ca]) в терминах вероятности
присоединения ионов к активным центрам
канала. Считалось, что электронный
переход может быть совершен в случае
присоединения более чем k
ионов Са2+
к активным центрам RyR-канала, состоящим
из z
активных мест присоединения, согласно
формуле (2.10). На рисунке 3.16 изображены
в логарифмических координатах графики
зависимостей интенсивностей электронных
переходов
![]() от cis[Ca] при различных значениях числаz
(мест присоединения активного центра
канала), и при различных значениях
концентрации Са2+
в cis-части, достаточной для заполнения
всех мест связывания на активном центре
(cis[Ca]max).
Как видно из графиков, насыщение
зависимости
от cis[Ca] при различных значениях числаz
(мест присоединения активного центра
канала), и при различных значениях
концентрации Са2+
в cis-части, достаточной для заполнения
всех мест связывания на активном центре
(cis[Ca]max).
Как видно из графиков, насыщение
зависимости
![]() достигается быстрее при малых значенияхz
и малых
значениях cis[Ca]max.
достигается быстрее при малых значенияхz
и малых
значениях cis[Ca]max.


В модели вводились переменные kab – минимальное число ионов Са2+, необходимых для связывания с активационным центром; kib – минимальное число ионов Са2+, необходимых для связывания с инктивационным центром для изменения состояния канала. Расчеты проводились при kab=4, kib=7, что соответствует предположительному числу ионов, необходимых для связывания с активационным и инактивационным центрами, соответственно. Максимальное число мест связывания активационного и инактивационного центров: z=50. Максимальная концентрация кальция: cis[Ca]max =4 мМ [20]. Зависимости интенсивностей электронных переходов представлены на рисунке 3.17.
3.4 Активация одиночного канала
На первом этапе проведения численных экспериментов проведено моделирование процесса активации канала при резком увеличении уровня цитозольного кальция. В начале эксперимента все каналы, входящие в статистический ансамбль, находятся в минимуме конформационного потенциала С, то есть в электронно и конформационно закрытом состоянии.
Для ансамбля, состоящего из 81 RyR-канала, вероятность пребывания в открытом состоянии в текущий момент времени равна:
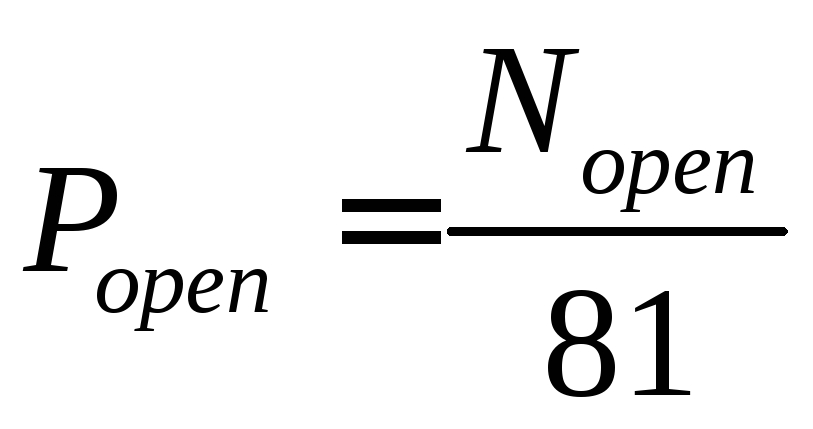 , (3.16)
, (3.16)
где
![]() –
число открытых каналов в текущий момент
времени.
–
число открытых каналов в текущий момент
времени.
На рисунке 3.18
представлены зависимости вероятности
пребывания канала в открытом состоянии
(Popen)
от времени при резком увеличении уровня
cis[Ca] от 0 до 1 мкМ в момент времени t=0.
Данные результаты получены из численных
экспериментов при различных значениях
интенсивности электронной активации
канала
![]() .
Этот параметр варьировался с целью
определения влияния вероятности
электронных переходов на длительность
процесса открытия RyR-канала.
.
Этот параметр варьировался с целью
определения влияния вероятности
электронных переходов на длительность
процесса открытия RyR-канала.

Анализируя графики,
можно сделать вывод, что зависимость
![]() носит экспоненциальный характер, причем
скорость увеличенияPopen
зависит от
носит экспоненциальный характер, причем
скорость увеличенияPopen
зависит от
![]() .
.
Для всех физических, биологических и химических характеристик, изменяющихся во времени экспоненциально, вводится понятие постоянной времени τ [117]. Физический смысл этого параметра следующий: он соответствует времени, за которое значение исследуемой величины увеличивается ровно в е раз, то есть τ характеризует скорость изменения исследуемой временной зависимости.
При изучении
процесса открытия одиночного канала
введена постоянная времени открытия
![]() .
Эта величина соответствует моменту
времениt,
при котором Popen
достигает
уровня:
.
Эта величина соответствует моменту
времениt,
при котором Popen
достигает
уровня:
![]() =
=![]() .
(3.17)
.
(3.17)
На рисунке 3.18
величина вероятности пребывания в
открытом состоянии оценивается
как![]() (пунктирная
линия).
(пунктирная
линия).
Зависимость
значения τopening
от интенсивности электронных переходов
![]() представлена на рисунке 3.19.
представлена на рисунке 3.19.

Из рисунка видно,
что при
![]() величина τopening
сначала
уменьшается, а затем достигает некоторого
стационарного значения 1.22 мс. Это
значение соответствует длительности
медленной конформационной релаксации
RyR-канала в открытое состояние.
величина τopening
сначала
уменьшается, а затем достигает некоторого
стационарного значения 1.22 мс. Это
значение соответствует длительности
медленной конформационной релаксации
RyR-канала в открытое состояние.
В ряде работ процесс активации RyR-канала при повышении уровня cis[Ca] в липидном бислое исследован экспериментально [24]. Для резкого повышения уровня cis[Ca] в основном применяются две методики. Первая из них заключается в высвобождении связанного Са2+ из сложных структур, находящихся в растворе, с помощью лазерного (или ультрафиолетового) флэш-фотолиза [25, 29, 106], во второй методике уровень cis[Ca] повышается механически, то есть увеличивается концентрация Са2+ вблизи канала в растворе [31, 119].
Результаты, полученные с помощью методики лазерного флэш-фотолиза, говорят о том, что постоянная времени активации канала составляет ~1 мс [25]. При механическом увеличении уровня кальция в растворе эта величина была немного больше и варьируется в интервале 2-20 мс [31].
Проведя анализ результатов численных экспериментов, можно сделать вывод, что они с хорошей степенью точности согласуются с этими экспериментальными данными.