

Глава 12
пищеварение
Организм человека в процессе жизнедеятельности расходует различные вещества и значительное количествоэнергии. Из внешней среды должны поступать вещества, восстанавливающие пластические и энергетические потребности организма. Длительное прекращение или явно недостаточное поступление питательных веществ приводит к нарушению гомеостаза и несовместимо с жизнью. Вместе с тем организм человека не способен ассимилировать белки, жиры, углеводы и ряд других веществ из пищи без предварительной обработки. Эту важнейшую функцию в организме осуществляет система пищеварения.
Пищеварением называется физическая и химическая переработка принятой пищи. В результате компоненты пищи, сохранив энергетическую и пластическую ценность, утрачивают видовую специфичность, но становятся доступными для усвоения организмом и включаются в нормальный обмен веществ.
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ГОЛОДА И НАСЫЩЕНИЯ
Голод как физиологическое состояние (в отличие от голодания как патологического процесса) служит выражением потребности организма в питательных веществах, которых он был лишен на некоторое время, что привело к снижению содержания этих веществ в депо и циркулирующей крови.
Субъективным проявлением голода являются неприятные ощущения: «жжения», чувство «сосания под ложечкой», тошнота, иногда — головокружение, головная боль,, чувство общей слабости.
11*
323
Субъективные и объективные проявления голода обусловлены возбуждением нейронов различных отделов и уровней ЦНС. Совокупность этих нейронов И. П. Павлов назвал пищевым центром. Его функциями являются формирование пищевого поведения, направленного на поиск и прием пищи, а также регуляция и функциональная интеграция органов пищеварительной системы.
Пищевой центр — это сложный гипоталамолимбико-ретикулокортикальный комплекс. Ведущим отделом, от которого распространяется активация всего пищевого центра, являются латеральные ядра гипоталамуса. При разрушении данных ядер возникает отказ от пищи (афагия), а их раздражение ведет к усиленному потреблению пищи (гиперфагия). Указанная часть пищевого центра обозначается как центр голода.
Разрушение вентромедиальных ядер гипоталамуса приводит к гиперфагии, а их раздражение — к афагии. На основании этого (и других фактов) считается, что данные ядра составляют центр насыщения.
Однако признание за гипоталамическими ядрами исключительной роли в формировании пищевого поведения, абсолютизирование их значения как пищевого центра неверно, гипоталамические ядра только часть этого центра. Нарушение пищевого поведения, хотя и не столь ярко, проявляется при поражении лимби-ческой системы, ретикулярной формации, передних отделов новой коры головного мозга. Электрофизиологические исследования подтверждают сложность строения и функций центрального комплекса, объединенного в пищевой центр в павловском его понимании.
Гипоталамические ядра пищевого центра возбуждаются или тормозятся в зависимости от состава крови, а также поступления разнообразных сигналов от различных периферических рецепторов.
Доказано, что у голодного животного, которому перелита кровь сытого животного, происходит угнетение рефлексов, направленных на добывание и прием пищи. Называют разные вещества, которые обеспечивают свойства «сытой» и «голодной» крови. В зависимости от вида этих веществ предложено несколько теорий.
Много сторонников у глюкостатической теории, согласно которой ощущение голода связано с понижением содержания глюкозы в крови. Полагают, что в гипоталамусе имеются глюкорецепторы, воспринимающие изменения содержания сахара в крови. Повышение уровня сахара в крови (например, при внутривенном введении глюкозы) снижало электрическую активность нейронов латерального ядра и несколько увеличивало ее в нейронах вентромедиального ядра гипоталамуса. При этом изменялся характер электрической активности коры передних отделов головного мозга: появлялись высокоамплитудные медленные электрические колебания, подобные тем, какие регистрируются у накормленных животных. Такой же эффект, как и введение глюкозы, оказывало вливание голодному животному крови сытого.
Однако имеются данные, противоречащие глюкостатической теории. Предложена аминоацидостатическая теория. По этой теории возбудимость нейронов пищевого центра определяется содержанием в крови аминокислот.
Липостатическая теория считает, что раздражителем гипоталамических центров является недостаток метаболитов, образующихся при мобилизации жира из жировых депо. Некоторые авторы утверждают, что пищевой центр стимулируется сигналами от жировых депо, когда из них высвобождается жир (липоневростатическая теория). Так называемая термостатическая теория предполагает угнетение пищевого центра вследствие повышения температуры омывающей его крови, что происходит во время приема пищи. Гидростатическая теория связывает чувство голода с водными ресурсами организма: снижение запаса воды в организме уменьшает потребление пищи.
Сейчас предложена и метаболическая теория, которая в известной мере соединяет все прежние. Согласно этой теории, промежуточные продукты цикла Кребса, образующиеся при расщеплении всех питательных веществ, циркулируя в крови, определяют степень пищевой возбудимости животного.
В последние годы из слизистой оболочки двенадцатиперстной кишки выделено вещество пептидной природы, которое при внутривенном введении вызывает снижение аппетита. Это вещество получило название аппетитрегулирующего вещества кишечного происхождения — арэнтерина. Угнетают аппетит и некоторые другие (например, холе- цистокинин-панкреозимин) интестинальные гормоны.
Не только изменение химического состава и свойств крови стимулирует или тормозит пищевой центр. В регуляции возбудимости пищевого центра существенная роль принадлежит афферентным влияниям от рецепторов пищеварительного тракта. Доказано, что наполнение желудка, в том числе раздувание в нем резинового баллона, тормозит пищевые реакции, а периодические сокращения свободного от пищи желудка вызывают ощущение голода. Афферентные влияния, поступающие по блуждающим и чревным нервам от пищеварительного тракта в ЦНС, способствуют формированию чувства голода или насыщения. Эти представления обозначают как локальную теорию голода. Несомненно, что в естественных условиях состояние пищевого центра определяется как составом крови, так и нервными сигналами от пищеварительных органов, депо питательных веществ, многочисленных интеро- и экстерорецепторов, от центров многих рефлексов.
Прием пищи вызывает противоположное голоду состояние насыщения. Оно возникает до того, как в кровь поступят продукты переваривания питательных веществ. Такое насыщение называют сенсорным (первичным) насыщением. Оно состоит в торможении пищевого центра и имеет сложную рефлекторную природу. Сенсорное насыщение сменяется обменным (вторичное, или истинное) насыщением, основным механизмом которого является поступление в кровь продуктов переваривания питательных веществ.
Известно избирательное влечение человека и животных к определенному виду пищи, чаще тому, который содержит недостающие организму вещества. Такое влечение к пище называют специфическим аппетитом. В его возникновении большое значение имеют изменения химического состава крови, которые воспринимаются интерорецепторами и гипоталамусом. В регуляции специфического аппетита велика роль вкусового анализатора и особенно его высшего отдела в коре больших полушарий головного мозга. Определенную роль играют также свойства содержимого желудка и тонкой кишки, рефлекторно влияющие на соответствующие центры, изменяя поведение и выбор той или иной пищи. Многие аспекты физиологии голода, насыщения и особенно специфического аппетита еще окончательно не выяснены.
Итак, состояние голода побуждает к поиску пищи, ее приему, а насыщение устраняет это состояние. Однако для того, чтобы питательные вещества поступили в кровь и были утилизированы организмом, пища должна пройти сложную механическую и химическую обработку в желудочно-кишечном тракте.
СУЩНОСТЬ ПИЩЕВАРЕНИЯ
И КЛАССИФИКАЦИЯ ПИЩЕВАРИТЕЛЬНЫХ ПРОЦЕССОВ
Пищеварение — сложный физиологический процесс, в ходе которого пища, поступившая в пищеварительный тракт, подвергается механическим и химическим превращениям, а содержащиеся в ней питательные вещества после деполимеризации всасываются в кровь и лимфу (рис. 165).
Физические изменения пищи заключаются в механической обработке, размельчении, набухании и растворении.
Химические изменения состоят из ряда последовательных реакций питательных веществ с компонентами секретов пищеварительных желез. В результате этих реакций происходит денатурация и последовательная деполимеризация — расщепление белков, жиров и углеводов под влиянием гидролитических ферментов (гидролазы) трех основных групп — соответственно протеаз, липаз и карбогидраз.
Ферменты вырабатываются секреторными клетками (гландулоцитами) пищеварительных желез и поступают в пищеварительный тракт в составе слюны, желудочного, поджелудочного и кишечного соков. Ферменты кишечного содержимого частично выделяются разрушенными слущившимися клетками эпителия слизистой оболочки кишки.
Пища-800—1000
г Вода-1.2-1.5 л
Рис.
165. Выделение секретов в полость
желудочно-кишечного тракта и
последовательность процессов в
пищеварительном конвейере.
Количество и соотношение ферментов в секретах пищеварительных желез соответствуют особенностям принятой пищи. Так, во время приема пищи, богатой белками, в секрете поджелудочной железы больше протеаз, углеводной — карбогидраз, а жирной пищи — липаз. В желудочно-кишечном тракте на пищу последовательно действуют секреты пищеварительных желез, содержащие ферменты; существует своего рода пищеварительный конвейер, итогом работы которого на каждом этапе являются все менее сложные химические соединения и, наконец, — мономеры. Продукты гидролиза белков (аминокислоты),
Рис. 166. Локализация гидролиза пищевых субстратов при разных типах пищеварения (по А. М. Уголсву).
а внеклеточное, дистантное пищеварение: б внутриклеточное пищеварение: в — пристеночное (контактное, мембранное) пищеварение, I — внеклеточная жидкость: 2 — внутриклеточная жидкость; 3 — внутриклеточная вакуоль: 4 — ядро; 5 - клеточная мембрана; 6 — ферменты.
жиров (моноглицериды, глицерин и жирные кислоты) и углеводов (моносахариды), лишенные видовой специфичности, но сохранившие свою энергетическую и пластическую ценность, всасываются в кровь и лимфу и используются клетками организма. Вода, минеральные соли и некоторые простые органические соединения пищи поступают в кровь в неизмененном виде.
В зависимости от происхождения гидролаз пищеварение делится на три типа: ауто- литическое,осуществляемое посредством ферментов, входящих в состав пищевых продуктов растительного или животного происхождения; симбионтное,при котором поставщиками гидролаз являются симбионты (бактерии, простейшие) данного макроорганизма; собственное— осуществляется ферментами, синтезируемыми в данном макроорганизме.
Так, у жвачных животных наряду с собственным пищеварением существует симбионтное пищеварение, при котором бактерии и простейшие расщепляют питательные вещества. У этих же видов животных гидролазы свежих кормов могут расщеплять собственные питательные вещества, т.е. имеются элементы аутолитического пищеварения. В период молочного вскармливания при недостаточно сформировавшемся собственном пищеварении у ребенка имеется аутолитическое пищеварение — в гидролизе питательных веществ молока принимают участие содержащиеся в нем гидролитические ферменты. Аутолитический тип пищеварения наименее изучен.
У высших животных ведущей формой пищеварения является собственное пищеварение, осуществляемое ферментами, продуцируемыми в организме, ассимилирующем пищу. Исключительное совершенство последнего у взрослого человека резко снизило роль других форм пищеварения.
Процессы пищеварения классифицируются также по их локализации. Выделяют внутриклеточноеи внеклеточное.Последнее в свою очередь делится на дистантное(полостное) и контактное(пристеночное, мембранное) (рис. 166).
Внутриклеточное пищеварение — это гидролиз питательных веществ, попавших внутрь клетки путем фагоцитоза или пиноцитоза. Питательные вещества гидролизуются клеточными (лизосомальными) ферментами либо в цитозоле, либо в пищеварительной вакуоли, на мембране которой фиксированы ферменты. В организме человека внутриклеточное пищеварение имеет место в лейкоцитах и клетках лимфоретикулогистио- цитарной системы. Роль внутриклеточного пищеварения в гидролизе питательных веществ относительно невелика.

О■ V <
. О-
«» а а* Q
' л. t$
р/л?Ь
^
о Р о-
66 }o6fi
-о
Ъ
-о
Л°6ъ*
а
frtf*»
Рис. 167. Взаимоотношении полостного и мембранного пищеварения (схема).
а — полость кишки без пищевых веществ; б - при их наличии. I — ферменты в полости кишки; 2 — микро- ворсинки; 3 ферменты на поверхности микровореннок, 4 — поры каемчатого эпителий; 5— микробы; 6—7 — пищевые вещества на различных стадиях гидролиза.
редственной близости от слизистой оболочки), ферменты поджелудочной железы — в полости тонкой кишки.
Контактное (пристеночное, мембранное) пищеварение осуществляется ферментами, фиксированными на клеточной мембране, на границе внеклеточной и внутриклеточной сред. При данной форме пищеварения гидролиз питательных веществ сопряжен с последующим транспортом в кровь и лимфу мономеров — продуктов гидролиза. Структуры, на которых фиксированы ферменты, представлены в тонкой кишке гликокаликсом (сете- видное образование из отростков мембраны микроворсинок). Именно здесь осуществляется пристеночное пищеварение, представляющее продолжение предшествовавшего ему полостного пищеварения в желудке и тонкой кишке (рис. 167). Иначе говоря, гидролиз питательных веществ первоначально производится в полости тонкой кишки ферментами поджелудочной железы, затем образовавшиеся олигомеры гидролизуются в зоне гликокаликса адсорбированными здесь ферментами поджелудочной железы, и, наконец, непосредственно у мембраны гидролиз образовавшихся димеров производят фиксирован-
и переносятся на мембраны их микроворсинок. Об этих процессах подробнее сказано при разборе кишечного пищеварения.
Таким образом, пищеварение состоит из преемственного действия на питательные вещества различных ферментов и всасывания продуктов гидролиза из зоны пристеночного пищеварения.
Основные этапы полостного пищеварения реализуются посредством ферментов, выделяемых в составе секретов пищеварительных желез, т. е. обеспечиваются секреторнойфункцией пищеварительного тракта. Указанные железы, кроме гидролаз, выделяют и другие вещества, создающие оптимальный рН для действия соответствующих ферментов, активирующие или ингибирующие их, вызывающие денатурацию пищевых белков.
Другой функцией пищеварительного тракта, также направленной на достижение эффективного гидролиза питательных веществ, является двигательная, или моторная. Она осуществляется мышечными волокнами пищеварительного аппарата и обеспечивает жевание, глотание, перемешивание пищи с пищеварительными секретами, передвижение пищи и непереваренных ее остатков по пищеварительному тракту с задержкой в том или ином отделе и порционной загрузкой его следующего отдела, а также выделение из организма непереваренных остатков пищи, некоторых продуктов его деятельности и бактерий. Моторика желудочно-кишечного тракта играет существенную роль в выведении в его полость пищеварительных секретов (открытие, закрытие сфинктеров протоков, их сокращения, сокращение желчного пузыря), в формировании складок слизистых оболочек, смене у их поверхности содержимого желудочно-кишечного тракта, сокращение ворсинок кишечника.
Еще одна функция пищеварительной системы — всасываниеиз полости желудка и кишечника продуктов гидролиза питательных веществ и самих секретов, их низкомолекулярных компонентов (вода, соли) и некоторых других веществ (витамины).
Органы желудочно-кишечного тракта осуществляют и непищеварительные функции. Прин-имают участие в обеспечении гомеостаза организма, выводя в составе секретов пищеварительных желез многие продукты обмена веществ (мочевина, желчные пигменты и др.) и вещества экзогенного происхождения, в том числе лекарственные. Данная функция называется экскреторной.Она отражает участие желудочно-кишечного тракта в межуточном обмене веществ. В регуляции обмена веществ всего организма принимают участие гормоны желудочно-кишечного тракта, образуемые огромным количеством расположенных в нем эндокринных клеток (диффузная эндокринная система). Симбионты, находящиеся в желудочно-кишечном тракте, синтезируют ряд физиологически активных веществ, участвуют в формировании иммунных свойств организма.
ПРИНЦИПЫ РЕГУЛЯЦИИ ПРОЦЕССОВ ПИЩЕВАРЕНИЯ
Деятельность пищеварительной системы регулируется нервными и гуморальными механизмами.
Нервная регуляция пищеварительной функции осуществляется пищевым центром с помощью условных и безусловных рефлексов, эфферентные пути которых образованы симпатическими и парасимпатическими нервными волокнами. Рефлекторные дуги могут быть «длинными» — их замыкание осуществляется в центрах головного и спинного мозга и «короткими», замыкающимися в периферических внеорганных (экстрамуральных) или внутриорганных (интрамуральных) ганглиях вегетативной нервной системы.
Вид и запах пищи, время и обстановка ее приема возбуждают пищеварительные железы условнорефлекторным путем. Чем с большим аппетитом поедается пища, тем секреция в этот период выше, но она может быть заторможена другими раздражителями. Прием пищи, раздражая рецепторы полости рта, вызывает безусловные рефлексы, усиливающие сокоотделение пищеварительных желез. Подобного типа рефлекторные влияния особенно выражены в верхней части пищеварительного тракта. По мере удаления от нее участие истинных рефлексов в регуляции пищеварительной функции уменьшается. Так,
Рис.
168.
Гормоны, вырабатываемые органами
же^удочио-кишечного
тракта, и места их образования.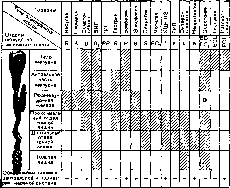
в наибольшей мере выражены рефлекторные влияния на слюнные железы, несколько меньше — на желудочные, еще меньше — на поджелудочную железу.
С уменьшением значения рефлекторных механизмов регуляции повышается значение гуморальных механизмов, особенно гормонов, образующихся в специальных эндокринных клетках слизистой оболочки желудка, двенадцатиперстной и тощей кишки, в поджелудочной железе. Эти гормоны названы гастроинтестинальными. В тонком и толстом отделах кишечника особенно велика роль локальных механизмов регуляции — местное механическое и химическое раздражение повышает активность кишки в месте действия раздражителей.
Таким образом, существует градиент распределения нервных и гуморальных регуля- торных механизмов в пищеварительном тракте, но регулировать деятельность одного и того же органа могут несколько механизмов. Например, секреция желудочного сока изменяется истинными рефлексами, гастроинтестинальными гормонами и локальными нейро-гуморальными механизмами.
Местные механические и химические раздражения влияют как путем периферических рефлексов, так и через гормоны пищеварительного тракта. Последние могут высвобождаться из эндокринных клеток и под действием химических веществ, которые влияют на эти клетки, диффундируя из полости желудка и кишечника. Химическими стимуляторами нервных окончаний в желудочно-кишечном тракте являются кислоты и щелочи (ЁГ- и ОН—ионы), продукты гидролиза питательных веществ. Поступая в кровь, эти продукты приносятся ее током к пищеварительным железам и возбуждают их непосредственно или через другие активные посредники, например биогенные амины. Некоторые из них, такие, как гистамин и серотонин, играют важную роль гуморальных регуляторов органов пищеварения.
Особенно велика роль в гуморальной регуляции деятельности органов пищеварения гастроинтестинальных гормонов. Некоторые из этих гормонов получены в чистом виде,
созданы их синтетические аналоги. Они находят широкое применение в клинико-диагностической и лечебной практике. Гастроинтестинальные гормоны относятся к числу пептидов и синтезируются специальными клетками слизистой оболочки желудка, кишечника и поджелудочной железы (рис. 168). Предполагается существование и других гормонов (гастрон, бульбогастрон, дуокринин, энтерогастрон, вилликинин и др.), но они не выделены и их химическая характеристика либо неполная, либо отсутствует.
Гастроинтестинальные гормоны оказывают множественные воздействия на функцию желудочно-кишечного тракта, некоторых других систем и обмен веществ всего организма. Эти гормоны влияют на секрецию воды, электролитов и ферментов, моторную активность желудочно-кишечного тракта, его сфинктеры; на всасывание воды, электролитов и питательных веществ, на пролиферативную активность слизистой оболочки и пищеварительных желез, на функциональную активность эндокринных клеток желудоч- но-кишечного тракта и некоторых эндокринных желез, на деятельность сердечно-сосуди- стой системы (табл. 16).
Таблица 16
Гормоны
Эффекты (наиболее выраженные выделены)
Гастрин
Секретин
ХЦК-ПЗ (холецистокинин-панкрео- зимин)
ЖИП (желудочный иигибирующий пептид)
ВИП (вазоактивный иитестиналь- ный пептид)
Мотилин
ПП (панкреатический полипептид)
Соматостатин Бомбезин
Энкефалич Нейротензни Вещество П (Р) Химоденнн Вилликинин
Усиление секреции желудка и поджелудочной железы, гипертрофия слизистой оболочки желудка, усиление моторики желудка, тонкой кишки и желчного пузыря Увеличение секреции бикарбонатов поджелудочной железой,
торможение секреции соляной кислоты в желудке Усиление сокращения желчного пузыря и желчевыделеиия, секреции ферментов поджелудочной железой, торможение секреции соляной кислоты в желудке, усиление н нем секреции пепсина, усиление моторики тонкой кишки Глюкозозавнсимое усиление высвобождения поджелудочной железой инсулина, торможение секреции и моторики желудка, торможение высвобождения гасгрина Расслабление гладких мышц кровеносных сосудов (снижение артериального давления), желчного пузыря, сфинктеров, торможение секреции желудка Усиление моторики желудка и тонкой кишки, усиление секреции пепсина желудком, синтеза белка Антагонист холецистокинин-панкреоэимнна, усиливает пролиферацию слизистой оболочки тонкой кишки, поджелудочной железы и печени, участвует в регуляции обмена углеводов и липидов
Торможение высвобождения желудочно-кишечных гормонов
и секреции желез желудка Стимуляция желудочной секреции через высвобождение гастрина, усиление сокращений желчного пузыря и выделение ферментов поджелудочной железой через высвобождение ХЦК-ПЗ, усиление высвобождения энтероглюкагона, нейротензина и ИМ Торможение секреции ферментов поджелудочной железой,
усиление высвобождения гастрина. Торможение секреции соляной кислоты железами желудка,
усиление высвобождения глюкагона Усиление моторики кишечника, слюноотделения, торможение
высвобождения инсулина и всасывания натрия Стимуляция секреции поджелудочной железой фермента
химотрнпсиногена Усиление моторики ворсинок тонкой кишки
Большинство гормонов обладает широкими спектрами действия н высвобождается из соответствующих клеток под влиянием нескольких факторов.
Некоторые гастроинтестинальные пептид-гормоны (гастрин, холецистокинин-панк- реозимин, ВИЛ, вещество П, энкефалин, нейротензин, бомбезиноподобный пептид) обнаружены в различных структурах мозга. Полагают, что это' результат эмбрионального происхождения эндокринных клеток из нейроэктодермы или .прилегающей к ней ткани. Значение этих веществ в ЦНС точно не установлено. Возможно, что данные пептид-гор- моны выполняют в мозге медиаторную функцию. Доказанной она считается лишь для некоторых пептидов (ВИП, соматостатин, энкефалин, вещество П, бомбезин), высвобождающихся в окончаниях ряда вегетативных нервных волокон, иннервирующих желудоч- но-кишечный тракт. Такие волокна названы пептидергическими (по аналогии с адрен- и холинергическими).
Перечисленные пептиды могут продуцироваться в соме нейронов, транспортироваться по аксону и высвобождаться при деполяризации его окончания. Здесь он оказывает (как нейромедиатор) локальное действие и быстро разрушается. Пептид может синтезироваться и в эндокринных клетках, высвобождаться из.них под влиянием ряда факторов и оказывать локальное влияние, диффундируя через интерстициальную жидкость от эндокринной клетки к клетке-мишени. Такое явление называется паракринией. К числу паракринных пептидов относят, например, ВИП, субстанцию П, соматостатин. И, наконец, пептиды, высвобождаясь в кровоток, циркулируют вместе с кровью в организме и действуют как типичные гормоны. Время их действия ограничивается несколькими минутами, после чего гормоны разрушаются в почках и печени и выводятся из организма. Для поддержания определенной концентрации пептид-гормонов в крови они должны непрерывно выделяться в кровоток клетками-продуцентами.
Действие интестинальных гормонов (энтеринов) не ограничивается пищеварительной системой.
Так, гормональные факторы двенадцатиперстной кишки увеличивают общий энергетический обмен, снижают аппетит, изменяют активность желез внутренней секреции и т. д. Таким образом, интестинальным гормонам принадлежит большая роль не только в регуляции деятельности органов пищеварения, но и обмена веществ организма в целом.
Секреторные и мышечные клетки желудочно-кишечного тракта изменяют уровень функциональной активности в зависимости от рефлекторных интеро- и экстерорецептор- ньгх и гуморальных влияний, а также в зависимости от уровня кровоснабжения (рис. 169). Механизмы регуляции взаимосвязаны и контролируют ход пищеварительного процесса на протяжении всего желудочно-кишечного тракта.
Эфферентные нервные и гормональные воздействия на органы пищеварения вызывают, по И. П. Павлову, три типа эффектов: функциональные, сосудодеигателъныеитрофические.Первый состоит в изменении той или иной функциональной активности клеток, органа или системы. Второй тип эффектов — изменение уровня их кровоснабжения, приведение его в соответствие с уровнем функциональной активности органа. Большинство нервных и гормональных стимуляторов органов пищеварения усиливает их кровоснабжение, что является важным условием поддержания высокой функциональной активности данных органов. Третий тип эффектов включает в себя разнообразные изменения трофики органов пищеварения: процессы синтеза в них пищеварительных секретов, изменения числа секреторных клеток в пищеварительных железах и т. д. Например, гастрин увеличивает число париетальных клеток в железах желудка; холецистокинин- панкреозимин - гландулоцитов в ацинусах поджелудочной железы.
И. П. Павлов основал учение о фазах секреции пищеварительных желез. По механизму изменения секреции пищеварительных желез выделяют две фазы: сложнореф- лекгорную и нервно-химическую. Первая осуществляется с помощью условных и безусловных рефлексов, вторая — нейрогуморальными механизмами. Деление на эти две фазы в значительной мере условно, так как нервные и гуморальные механизмы взаимосвязаны. Секреция начинается с «мозговой» фазы и осуществляется по типу условного (вид, запах пищи и т.д.), безусловного (раздражение рецепторов полости рта и пищевода) рефлексов.
Эксторорвцепторы
I Ч
ЦНС
Рис.
169. Механизмы регуляции секреторной и
моторной деятельности органон
пищеварения.
Внешний
По маету афферентных нейро-гуморальных воздействий -
„Мозговая фаза" С&сг аец*пт| С рецепт, полости!