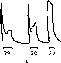Рефрактерный период
Воемя. мс 5
Рис.21. Изменения возбудимости нервного волокна в различные фазы развития потенциала действия и следовых изменений мембранного потенциала.
Рис.
22. Изменения потенциалов действия
нервного волокна в фазу относительной
рефрактерности.
На
нервное волокно многократно наносятся
два сильных стимула, разделенных
различными интервалами.
Супернормальный
период![]()
_
Субнормальный^ период
снижению калиевой проводимости. Это период так называемой относительной рефрактерности. В данную фазу возбудимость постепенно возрастает.
В нервных волокнах длительность фазы относительной рефрактерности составляет 5—10 мс. При наличии следовой деполяризации фаза относительной рефрактерности сменяется фазой повышенной возбудимости («су пер нормальности»). В этот период пороговый потенциал AV и соответственно порог раздражения снижены по сравнению с исходными значениями, поскольку мембранный потенциал ближе к критической величине, чем в состоянии покоя. В быстрых двигательных волокнах теплокровных животных фаза следового повышения возбудимости продолжается до 30 мс (рис. 22).
Следовая гиперполяризация, напротив, сопровождается снижением возбудимости. По своему механизму это снижение возбудимости сходно с тем, которое имеет место при анэлектротоне: AV увеличен за счет удаления мембранного потенциала Еот критической величины Ек.Сходство это, однако, неполное: при анэлектротоне gK низко, а во время следовой гиперполяризацип gK увеличено, что также повышает порог раздражения.
МЕХАНИЗМЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ
Проведение возбуждения вдоль нервных и мышечных волокон осуществляется при помощи так называемых местных токов, возникающих между возбужденным (деполяризованным) и покоящимися (нормально поляризованными) участками волокна. Распространение местных токов по длине волокна определяется его кабельными свойствами. Направление местного тока, показанного на рис. 23 таково, что он деполяризует соседний с активным (А) покоящийся (В) участок мембраны. Деполяризация эта быстро достигает критического уровня и порождает потенциал действия, который в свою очередь активирует соседний покоящийся участок. Благодаря такому эстафетному механизму возбуждение распространяется вдоль всего волокна. В мышечных и без-
А — ^Б
♦
+
+ ± ^^t
j^t+ttt+i
+
4 + -i-
+
Рис. 23. Принципиальный механизм распространения возбуждения с помощью местных токов (схема
-Jr
. А
"I М-J
АГ
^|Б +
+ +
+
+
+
++ ++++ +
■в
НЦ1
—
J
шт\\-
+
+ + + + + ~ ~
-+
+ + + + + - - +
+ + + + + + + +
Рис. 24. а расположение раздражающих (Р) и отводящих (А, Б) электродов для отведения двухфазного (I) и монофазного (II) потенциалов действия (схема); б — механизм возникновения двухфазного потенциала действия (схема).
мякотных нервных волокнах возбуждение осуществляется непрерывно «от точки к точке». Особенности проведения возбуждения по миелинизированным волокнам рассмотрены далее.
Распространение возбуждения в нерве или мышце можно зарегистрировать в эксперименте, если к двум точкам — А и Б (рис. 24, а) приложить отводящие электроды, связанные с регистрирующей аппаратурой, а к другой точке (Р) — раздражающие электроды. При нанесении электрического стимула на экране осциллографа регистрируется двухфазное колебание потенциала.
Схема на рис. 24, б разъясняет механизм возникновения этих двух противоположно направленных отклонений потенциала. В состоянии покоя все участки наружной поверхности возбудимой мембраны заряжены электроположительно по отношению к ее внутренней поверхности. Когда волна возбуждения проходит через участок под электродом, ближайшим к месту раздражения, наружная поверхность мембраны в этом участке становится электроотрицательной по отношению к точке Б. Это вызывает отклонение луча осциллографа вверх. Когда волна возбуждения покидает этот участок, луч возвращается в исходное положение. Затем возбуждение достигает участка под вторым электродом Б; этот участок в свою очередь становится электроотрицательным по отношению к точке А, а луч осциллографа отклоняется вниз. Если участок нерва под дальним электродом Б сделать невозбудимым при воздействии какого-либо агента, например новокаина, либо нарушить проведение возбуждения между участками А и Б, вторая фаза колебаний потенциала исчезает и регистрируемый потенциал действия становится однофазным.
+
+ + +-*- + + ^ 6
через этот участок полностью прекратится. Однако оно тотчас восстанавливается, если два разобщенных неэлектролитом участка волокна соединить металлическим проводником.
Скорость проведения зависит не только от сопротивления окружающей волокно среды, но и от внутреннего сопротивления волокна (т. е. сопротивления аксоплазмы на единицу длины). С увеличением диаметра волокна это сопротивление падает, поэтому скорость проведения возрастает. При одном и том же диаметре волокна скорость зависит главным образом от величины так называемого фактора надежности, который представляет собой отношение:
Амилитуда потенциала действия, мВ Порог деполяризации, мВ
Чем фактор надежности больше, тем скорость проведения выше и наоборот.
В нервных волокнах фактор надежности обычно 5—6. Это означает, что для покоящихся участков мембраны распространяющийся потенциал действия является сильным раздражителем, обладающим большим избытком мощности. Поэтому, для того чтобы заблокировать проведение нервного импульса, необходимо либо сильно повысить величину порога деполяризации нервного волокна, либо очень значительно снизить амплитуду его потенциала действия. Местные обезболивающие препараты (новокаин, кокаин, дикаин), применяемые в медицинской практике, вызывают оба этих изменения одновременно.
ОБМЕН ВЕЩЕСТВ ПРИ ВОЗБУЖДЕНИИ
При возникновении и проведении возбуждения в нервных клетках и мышечных волокнах проводит усиление обмена веществ. Это проявляется как рядом биохимических изменений, происходя- цих в мембране и цитоплазме клеток, так и усилением их теплопродукции.
Биохимическими и гистохимическими методами исследования установлено, что при возбуждении габлюдается усиление распада в клетках богатых энергией фосфорных соединений — аденозинтри- [юсфата (АТФ) и креатинфосфата (КФ); процессов распада и синтеза углеводов, белков и липидов; жислительных процессов, приводящих в сочетании с гликолизом к ресинтезу АТФ и КФ; происходят ;интез и разрушение медиаторов, например ацетилхолина и норадреналина; усиление синтеза РНК I белков.
МАКСИМАЛЬНЫЙ РИТМ ИМПУЛЬСАЦИИ
В естественных условиях существования организма по нервным волокнам проходят ie одиночные потенциалы действия, а серии импульсов, следующих друг за другом с раз- шчными интервалами. В двигательных нервных волокнах при произвольных движениях шстота импульсации обычно не превышает 50 в секунду, т. е. межимпульсный интервал доставляет около. 200 мс. При таком большом интервале все восстановительные процессы, развивающиеся после окончания потенциала действия (реактивация натриевых кана- юв, восстановление исходной натриевой проводимости, «откачка» из цитоплазмы ионов \Га+ и возвращение внутрь волокна ионов К+и т. д.), успевают полностью закончиться. Зднако в чувствительных нервных волокнах (например, в слуховом или зрительном нерве) фи сильном раздражении в начальный момент частота разряда может достигать 1000 и юлее импульсов в секунду при длительности абсолютной рефрактерной фазы 0,5—0,7 мс. Тодобные высокочастотные разряды импульсов появляются при возбуждении и в некото- )ых нервных клетках, например в клетках Реншоу спинного мозга.
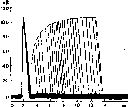
Н. Е. Введенский первый обратил внимание на разную способность возбудимых образований воспроизводить высокие ритмы раздражений. Максимальное число потенциалов действия («максимальный ритм»), которое способно возбудимое образование генерировать в 1 с в соответствии и с ритмом раздражения, Н. Е. Введенский предложил в сачестве показателя «лабильности»ткани. В настоящее время ясно, что максимальный
4J ч
Рис. 25. Стадии нарушения проведения через «парабиотический участою> нерва (по Н. Е. Введенскому).
а — соотношение меяеду силой ритмической стимуляции нормального нерва и высотой тетануса скелетной мышцы. Цифры означают расстояние меяеду катушками индукционного аппарата: чем ближе это расстояние— сильнее раздражение, тем больше частота нервных импульсов и соответственно выше тетанус; бив — то же после смазывания участка нерва 0,5 % раствором кокаина: б — «уравнительная фаза парабиоза»: раздражения разной силы (частоты) вызывают примерно одинаковый эффект; в — «парадоксальная фаза»: сильные (частые) стимулы дают меньший эффект, чем слабые (редкие).
(предельный) ритм импульсации нервных и мышечных волокон определяется скоростями процессов изменений ионной проводимости, лежащих в основе абсолютной и относительной рефрактерности.
Существуют воздействия, замедляющие реактивацию натриевых каналов и потому увеличивающие длительность фазы относительной рефрактерности. К ним относятся, например, местные анестетики. Поэтому участок нервного волокна, подвергнутый воздействию малых концентраций новокаина (или какого-либо другого местного анестетика), утрачивает способность проводить высокочастотные разряды импульсов, тогда как низкочастотные разряды еще продолжают проходить.
При высокочастотной стимуляции происходит либо трансформация ритма (блокируется каждый второй потенциал действия), либо (при очень частой стимуляции) проходит только первый потенциал действия, а остальные оказываются заблокированными. Объясняется это тем, что при частой стимуляции потенциалы действия, приходящие в альтерированный участок, углубляют инактивацию натриевых каналов, вызванную анестетиком'.
Сходным образом влияют на процесс реактивации повышение концентрации ионов К+в окружающей нервные волокна жидкости и некоторые другие химические агенты.
Н. Е. Введенский (1901) впервые обнаружил нарушение способности нерва проводить высокочастотные разряды импульсов при воздействии на нерв разных химических агентов (рис. 25). Он правильно усмотрел определенное сходство между состоянием, в котором находится нервное волокно при его альтерации химическими агентами, и состоянием рефрактерности, сопровождающей нормальный потенциал действия: и в том и вдругом случае, как это теперь установлено, происходит инактивация натриевых каналов.