

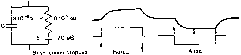
Рис.
13. Простейшая электрическая схема,
воспроизводящая электрические свойства
мембраны (а) и изменения мембранного
потенциала под катодом и анодом
постоянного тока подпороговой силы
(б).
а:
С — емкость мембраны, R —
сопротивление, Е — электродвижущая
сила мембраны в покое (потенциал покоя).
Приведены средние значенияR,
С и Е для мотонейрона, б — деполяризация
мембраны (1) под катодом и гиперполяризация
(2) под анодом при прохождении через
нервное волокно слабого подпорогового
тока.Наружная сторона Потенциал
или «электротоническими»,изменениями мембранного потенциала. В чистом виде последние могут быть зарегистрированы в условиях полной блокады ионных каналов химическими агентами. Различают кат-и анэлектротоническиеизменения потенциала, развивающиеся в области приложения соответственно катода и анода постоянного тока.
Критический уровень деполяризации
Регистрация изменений мембранного потенциала при внутриклеточном раздражении нервного или мышечного волокна показала, что потенциал действия возникает в тот момент, когда деполяризация мембраны достигает критического уровня. Этот критический уровень деполяризациине зависит от характера примененного стимула, расстояния между электродами и т. п., а определяется исключительно свойствами самой мембраны.
Рис.
14. Изменение мембранного потенциала
до критического уровня деполяризации
мембраны при действии раздражающего
тока разной силы и длительности.
Критический
уровень показан пунктиром. Внизу —
раздражающие стимулы, при воздействии
которых получены ответы А, Б н В.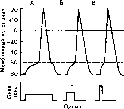
Локальный ответ
В механизме критической деполяризации мембраны наряду с пассивными существенную роль играют активные подпороговые изменения мембранного потенциала, проявляющиеся в форме так называемого локального ответа.
В
Рис.
15. Локальный ответ нервного волокна.
А, Б, В -
изменения мембранного потенциала
нервного волокна, вызываемые действием
подпоро- гиноги гока короткой длительности.
На кривых Б и В к пассивной деполяризации
мембраны присоединяется и активная
подпороговая деполяризация в форме
локальнога ответа. Локальный ответ
отделен от пассивных изменений
потенциала пунктирной линией. При
пороговой силе гока (Г) локальный
ответ перерастает в потенциал действия
(его вершина на рисунке не показана).

С
F
D
Реобаза А
О
Время
Рис. 16. Кривая силы — длительности. Объяснение в тексте.
Первые признаки локального ответа появляются при действии стимулов, составляющих 50—75 % от пороговой величины. По мере дальнейшего усиления раздражающего тока локальный ответ увеличивается, и в момент, когда деполяризация мембраны, обусловленная суммой катэлектротонического потенциала и локального ответа, достигает критического уровня, возникает потенциал действия (рис. 15).
Локальный ответ, так же как и потенциал действия, обусловлен повышением натриевой проницаемости мембраны. Однако при подпороговом стимуле это начальное повышение натриевой проницаемости недостаточно велико, чтобы вызвать быструю регенеративную деполяризацию мембраны. Развитие деполяризации тормозится процессами инактивации натриевых и активации калиевых каналов. Поэтому рост локального ответа приостанавливается, а затем происходит реполяризация мембраны. Амплитуда локального ответа увеличивается по мере приближения силы стимула к порогу, и при достижении последнего локальный ответ перерастает в потенциал действия, поскольку скорость увеличения натриевой проницаемости мембраны начинает превышать скорость роста калиевой проницаемости.
Зависимость пороговой силы раздражителя от его длительности
Пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности. Особенно четко эта зависимость проявляется при использовании в качестве раздражителя прямоугольных импульсов постоянного тока.
Представленная на рис. 16 кривая называется кривой силы — длительности, или силы — времени. Она была изучена при исследовании различных нервов и мышц Гоорве- гом (1892), Вейсом (1901) и Лапиком (1909).
По этой кривой прежде всего можно судить о том, что ток ниже некоторой минимальной силы или напряжения не вызывает возбуждения, как бы длительно он ни действовал. Минимальная сила постоянного тока, способная вызвать возбуждение (порог раздражения), названа Лапиком реобазой(ординатаOA). Наименьшее время (отрезок ОС), в течение которого должен действовать раздражающий стимул, величиной в одну реобазу называют полезным временем.Слово «полезное» здесь применено с целью подчеркнуть, что дальнейшее увеличение длительности действия тока не имеет значения (бесполезно) для возникновения потенциала действия.
Усиление тока приводит к укорочению минимального времени раздражения, но не беспредельно. Как видно на рис. 16, при очень коротких стимулах кривая силы — времени становится параллельной оси ординат. Это означает, что при таких кратковременных раздражениях возбуждения не возникает, как бы ни была велика сила раздражителя. Поэтому, кроме полезного времени, в качестве времени константы раздраже
ния Лапик ввел понятие «хронаксия». Хронаксия - это время, в течение которого должен действовать ток удвоенной реобазы, чтобы вызвать возбуждение.
мВ
•00
20
30
мс
Явление аккомодации
Пороговая сила тока увеличивается при уменьшении крутизны его нарастания, а при некоторой минимальной крутизне ответы на раздражение исчезают. Это явление принято обозначать термином «аккомодация». На рис. 17, а показаны изменения критического уровня деполяризации и амплитуды потенциала действия при раздражении одиночного нервного волокна лягушки линейно нарастающими токами различной крутизны. Уменьшение последней приводит к повышению критического уровня деполяризации (примерно на 20 % от исходной величины) и снижению амплитуды потенциалов
—-
XIU
J
J!r—
а з
jy!
ju_ JUJUL
Jfc
J/y/U

действия. При снижении крутизны до некоторого минимального уровня («минимальный градиент», или «критический наклон») потенциал действия не возникает. Величина этого «минимального градиента», выраженного в единицах реобаза в секунду, принята в качестве меры скорости аккомодации.
В основе аккомодации лежат инактивация натриевой и повышение калиевой проводимостей, развивающиеся во время медленно нарастающей деполяризации мембраны. Аккомодация различных нервных волокон варьирует в широких пределах, но у двигательных нервных волокон скорость аккомодации, как правило, значительно выше, чем у чувствительных волокон.
Повторные ответы
Многие нервные волокна обладают способностью отвечать несколькими потенциалами действия на включение постоянного деполяризующего тока. Эта способность к повторным ответам особенно хорошо выражена у сенсорных волокон, для которых длительная деполяризация мембраны рецептора является естественным раздражителем. Примеры повторных ответов на постоянный ток представлены на рис. 17, б. Как правило, пороговая сила тока, необходимая для возникновения повторных ответов, выше, чем для инициации одиночного потенциала действия. На рисунке видно, что увеличение силы деполяризующего тока до определенной величины обусловливает возрастание частоты импульсов и увеличение их числа (1—4). Однако при дальнейшем повышении силы тока частота импульсов уменьшается и в конечном итоге возникает только одиночный потенциал действия (5—8).
При постоянной силе тока длина межимпульсных интервалов в повторном ответе постепенно увеличивается. Это явление получило название адаптации.В его основе лежит медленное повышение калиевой проводимости мембраны, связанное с активацией особых медленных калиевых каналов. Эти каналы найдены как в нервных волокнах, так и в нервных клетках, у которых способность к повторным ответам и явление адаптации, как правило, хорошо выражены.
Изменение критического уровня деполяризации
Было показано, что условием возникновения потенциала действия является критическая деполяризация мембраны. Если исходный потенциал мембраны перед нанесением раздражающего стимула обозначить £о, а критическую величину мембранного потенпиала Е^. то указанное условие порогового раздражения можно записать так: £о+Д1/= £,,, где ДК — пороговый потенциал, т. е. величина, на которую необходимо повысить внутренний потенциал мембраны для возникновения импульса.
Эта простая зависимость помогла понять причину известных со времени классических исследований Пфлюгера (1859) изменений возбудимости нервного волокна в области катода и анода постоянного тока. В области катода мембрана деполяризуется, т. е. значение Е0приближается к величине Ек, соответственно Д 1/уменыпается и, следовательно, возбудимость возрастает. В области анода, напротив, Ъо уменьшается (внутренний потенциал мембраны становится более отрицательным); теперь для критической деполяризации мембраны необходимо ее потенциал сместить на большую величинуW—возбудимость снижается (рис. 18).
При длительной деполяризации мембраны развиваются процессы, повышающие критический уровень деполяризации. Такими процессами являются инактивация натриевых каналов и активация калиевых. Рост Екпри данном значении Е0ведет к увеличению порогового потенциала, т.е. снижению возбудимости (рис. 19). Одновременно с увеличением порога происходит снижение потенциала действия — крутизна его нарастания и амплитуда падают. Такое снижение возбудимости нервного волокна при длительной и сильной деполяризации мембраны было впервые описано Вериго (1889) и получило название катодической депрессии.
100
MB
1_
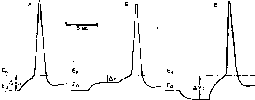
Рис. 18. Соотношения между исходным уровнем потенциала покоя (Ео), критическим уровнем деполяризации (Ек) и пороговым потенциалом(AV), в норме (А) и при электротоне (Б,В).
Б — катэлсктротоническое снижение порога AVi наступает в результате приближения потенциала покоя Ео к критическому уровню деполяризации Ек; В — анэлектротоническое повышение порога AV2 является следствием удаления исходного уровня потенциала покоя Е0от Ек. Нижняя линия — раздражающий и поляризующий токи: катодный ток — вверх, анодный ток — вниз.
Описанные явления демонстрируют наблюдающиеся в естественных условиях изменения возбудимости нервных клеток и пресинаптическйх нервных терминалей при действии на них медиаторов, вызывающих кратковременную или длительную деполяризацию мембраны. В частности, предполагают, что явление католической депрессии лежит в основе так называемого пресинаптического торможения, наблюдающегося в ЦНС.
ив
100
50
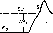
Ен
[2
1 ис
Рис. 19. Католическая депрессия Вериго.
I— ответ нормального нервноговолокна на пороговый раздражающий ток: 2 — ответ того же волокна через 10 смосле начала действиякатода по дно ро го но го постоянного гока, Е,:, Ек нAV — исходные величины потенциала покоя, критического уровня деполяризации и порогового потенциала. F.'h. F.\ и AVi - величины тех же параметров новремя католической депрессии.
Рис.
20. Возникновение
а иод но-
размыка тельного возбуждения.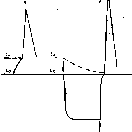
а — потенциал действия, возникающий под катодов при замыкании тока; б — потенциал действия, возникший иод анодом при размыкании сильного тока. Стрелкой вниз показано нключенне анодного тока, стрелкой вверх его выключение.
В заключение необходимо рассмотреть изменения возбудимости, наблюдающиеся при длительной гиперполяризации мембраны. Такая гиперполяризация (в нервном волокне она появляется в области приложения анода) приводит к снижению калиевой проницаемости и ослаблению исходной натриевой инактивации. Эти изменения ведут к снижению AV возросшего в начальный момент гиперполяризации мембраны, и увеличению амплитуды и крутизны нарастания потенциала действия. Если гиперполяризу- ющий ток прикладывается к мембране, которая предварительно была подвергнута воздействию избытка ионов или действию анестетиков, усиливающих исходную натриевую инактивацию (т. е. увеличивающих долю каналов, у которых инактивационные «ворота» закрыты), то во время длительной гиперполяризации АV может снизиться настолько, что при выключении тока, т. е. при возвращении мембранного потенциала к его исходной величине, возникает потенциал действия (рис. 20). Такое явление получило название анодно-размыкательного возбуждения. Предполагают, что в некоторых нервных клетках на подобном механизме основано возникновение потенциала действия после окончания тормозного гиперполяризационного потенциала в естественных условиях. По-видимому, у таких клеток инактивация натриевых каналов и активация калиевых сильно выражены при потенциале покоя в нормальной солевой среде.
Изложенные данные о влиянии деполяризации и гиперполяризации мембраны на ее возбудимость представлены в табл. 2.
|
Физиологические |
Деполяризация мембраны |
Гиперполяризация мембраны |
|
параметры |
(катэлектротон) |
(анэлектротон) |
|
Возбудимость |
Вначале повышение, затем пони |
Сначала понижение, затем относи |
|
|
жение |
те ттт,нпе ппитаптение |
|
Потенциал действия |
Понижение, прогрессирующее во |
Повышение, прогрессирующее во |
|
|
времени вплоть до полного |
времени |
|
|
vrnetennff |
|
|
Скоростьпроведения |
Вначале повышение, затем пони |
Вначале понижение вплоть до бло- |
|
|
жение rttttott, тто полного бттокя |
кя Сппи питтт,ном токе"* чятем |
|
|
|
постепенное рюеетянортение |
|
Натриевая |
Вначале повышение, приводящее |
Постепенное ослабление инактива |
|
мость |
к возникновениюпри пояпорого- |
ции. если оня имела место |
|
|
rom токе ттокя ттт,ного отиетя |
|
|
|
а при пороговом — потенциала |
|
|
|
лейглтшя -чятем ппетепенняя |
|
|
|
иняктивяттия |
|
|
Калиевая проницаемость |
Постепенное повышение |
Понижение, если она была по |
|
|
|
вышена |
ИЗМЕНЕНИЯ ВОЗБУДИМОСТИ ПРИ ВОЗБУЖДЕНИИ
Используя тестирующие стимулы в различные фазы развития потенциала действия, можно проследить временной ход изменений возбудимости, сопровождающих возбуждение. На рис. 21 видно, что во время развития локального ответа возбудимость повышается (мембранный потенциал приближается к критическому уровню деполяризации); во время пика потенциала действия мембрана утрачивает возбудимость, которая постепенно восстанавливается после окончания пика. Период полной невозбудимости получил название «фаза абсолютной рефрактерности». Она обусловлена практически полной инактивацией натриевых каналов и повышением калиевой проводимости. Реполяризация мембраны ведет к реактивации натриевых каналов и
Высоковольтный
потенциал
Положи
те,-ьныи следонои потенцийп