

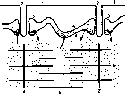
0.2 СРис. 28. Кривые двух одиночных сокращений н :u.U про найми го мышечного ммокна.
Оптическая регистрация. М — многрачма; Р от Mt'i кд раздражения: В н [метка иремени 0,2 с {по Ф П. Серкову).
Кривые одиночного сокращения изолированного мышечного волокна сходны по форме с кривыми одиночных сокращений целой мышцы (рис. 28). Отличием является меньшая длительность сокращения одиночного волокна но сравнению с целой мышцей.

/\л
115
35
2 &
20
Рис. 30. Сокращения икроножной мышцы при разной частоте Рис. 29. Суммация мышечных раздражения (но Е. Е. Жукову).
сокращений н ответ на 2 сти- Одиночное сок решение показано пунктиром Цифры над кривыми —
мула. Or метка времени 20 мс. частоты стимулов. При частотах 20- 33 н секунду - губчатый тета-
Объяснение в тексте. нус, при частоте 115 в секунду — гладкий тетанус.
Рис 31. Тетанус изолированного мышечного волокна (по Ф. II. СерковуЬ
а — -губчатый тетанус при частоте ратдрижения Гц: б - гладкий тетинус при частоте раздражения 35 Гц; М — мииграмма; Р - отметки раздражения; В — отметка времени 1 с.
Рис. 32. Одновременная запись сокращения (а) и 'электрической активности (б) скелетной мышцы кошки при чета ни чес ком раздражении нерва.

графической кривой вершина этого сокращения будет отделена от вершины первого запа- денисм (рис. 29, Ж — Г). Еели же второе раздражение действует, когда первое еще не дошло до своей вершины, то второе сокращение полностью сливается с первым, образуя единую суммированную вершину (рис. 29, А — В).
Рассмотрим суммацию в икроножной мышце лягушки. Продолжительность восходящей фазы ее сокращения примерно 0,05 с. Поэтому для воспроизведения на этой мышце первого типа суммации сокращений (неполная суммация) необходимо, чтобы интервал между первым и вторым раздражениями был больше 0,05 с, а для получения второго типа суммации (так называемая полная суммация) — меньше 0,05 с.
Как при полной, так и при неполной суммации сокращений потенциалы действия не суммируются.
Тетанус мышцы.Если на отдельное мышечное волокно или на всю мышцу действуют ритмические раздражения с такой частотой, что их эффекты суммируются, наступает сильное и длительное сокращение мышцы, называемое тетаническим сокращением, или тетанусом. Амплитуда его может быть в несколько раз больше величины максимального одиночного сокращения.
При относительно малой частоте раздражений наблюдается зубчатый тетанус,при большой частоте — гладкий тетанус(рис. 30, 31). При тетанусе сократительные ответы мышцы суммированы, а электрические ее реакции — потенциалы действия — не суммируются (рис. 32) и их частота соответствует частоте ритмического раздражения, вызвавшего тетанус.
После прекращения тетанического раздражения волокна полностью расслабляются, их исходная длина восстанавливается лишь по истечении некоторого времени. Это явление называется цоелстетанической, или остаточной, контрактурой.
Чем быстрее сокращаются и расслабляются волокна мышцы, тем чаще должны быть раздражения, чтобы вызвать тетанус.
Двигательные единицы
Чтобы познакомиться с тем, как функционирует мышца в условиях естественной деятельности организма, необходимо остановиться на строении и особенностях иннервации скелетной мышцы двигательным нервом.
Каждое двигательное нервное волокно является отростком нервной клетки — мотонейрона,расположенного в переднем роге спинного мозга или в двигательном ядре черепного нерва. В мышце двигательное волокно ветвится и иннервирует не одно, а целую группу мышечных волокон. Мотонейрон вместе с группой иннервируемых им мышечных волокон называется двигательной единицей.
Среднее количество мышечных волокон, входящих в состав двигательной единицы, в разных мышцах варьирует в широких пределах. Так, у человека в некоторых мышцах глазного яблока двигательные единицы содержат в среднем менее 10 мышечных волокон, в отдельных мышцах пальцев руки — 10—25. В отличие от этого в большинстве мышц туловища и конечностей на одно двигательное волокно приходится в среднем сотни мышечных волокон, а в камбаловидной мышце — 2000.
Когда по двигательному волокну к мышце приходит потенциал действия, мышечные волокна, входящие в одну двигательную единицу, возбуждаются почти одновременно. Возникающий при этом суммарный потенциал действия мышечных волокон двигательной единицы может быть зарегистрирован вколотым в мышцу электродом с малой отводящей поверхностью. Поскольку мотонейрон при естественном сокращении мышцы разряжается ритмически, электрическая активность двигательной единицы имеет в записи вид частокола (рис. 33). У здорового человека в расслабленной мышце (т. е. в состоянии полного покоя) электрическая активность в мышце почти отсутствует. При небольшом напряжении мышцы, например, связанном с поддержанием позы, двигательные единицы разряжаются с частотой 5—10 импульсов в секунду (имп/с), при увеличении силы сокращения частота повышается до 20—30, лишь при максимальном напряжении мышцы она может достигать 50 имп/с или несколько более. Исследование частоты разрядов двигательных единиц позволило установить, что в естественных условиях сокращения мышц мотонейроны разряжаются со сравнительно низкой частотой. Более высокие частоты зарегистрированы только при исследовании двигательных единиц мышц глазного яблока (150 имп/с и более).
Функциональная дифференциация двигательных единиц
В скелетных мышцах теплокровных животных и человека различают быстрые и медленные двигательные единицы, состоящие соответственно из быстрых и медленных мышечных волокон. Длительность сокращения медленных двигательных единиц может быть 100 мс и более, быстрых —10—30 мс. Существуют мышцы, состоящие преимущественно из быстрых двигательных единиц (например, мышцы глазного яблока), и мышцы, в которых преобладают медленные двигательные единицы (например, камбало- видная мышца). Такие мышцы часто называют соответственно быстрыми и медленными. Большинство мышц смешанные, состоят как из быстрых, так и из медленных двигательных единиц, а также переходных форм между ними.
шш
0,6
Рис.
33. Электрическая активность отдельной
моторной единицы (а) и целой мышцы (б)
у человека (по Ф. Бухталю).
Сопоставление частоты разрядов двигательных единиц с частотой, при которой может образоваться гладкий тетанус, позволяет сделать вывод, что в естественных условиях гладкий тетанус может наблюдаться только при очень высокой частоте. Обычным режимом естественного сокращения является зубчатый тетанус или даже ряд последовательных одиночных сокращений двигательной единицы. Тем не менее это не отражается на сокращении целой мышцы; оно, как правило, бывает слитным, напоминающим гладкий тетанус. Причина этого — асинхронность разрядов мотонейронов, а следовательно, и мышечной части двигательных единиц. При отведении игольчатым электродом потенциалов действия одновременно нескольких активных двигательных единиц видна асинхронность их импульсации. В случае электрической активности целой мышцы сложение (интерференция) потенциалов действия многих двигательных единиц дает сложную картину колебаний потенциала, в которой уже не удается различать потенциал каждой из них, а общая частота колебаний существенно превышает частоту разрядов каждой из активных двигательных единиц.
Скорость сокращения двигательных единиц коррелирует и с другими их свойствами. Медленные двигательные единицы, как правило, содержат меньше мышечных волокон и, следовательно, при сокращении развивают меньшую силу. Количество мышечных волокон и развиваемая ими суммарная сила в двигательных единицах одной мышцы могут различаться более чем на порядок. Не менее важно другое различие медленных и быстрых двигательных единиц — устойчивость к утомлению. Медленные двигательные единицы могут работать без утомления гораздо дольше, чем быстрые, что объясняется особенностями их обмена.
Со свойствами мышечных волокон двигательной единицы коррелируют и свойства иннервирующего ее мотонейрона: при естественном напряжении мышцы мотонейроньг медленных двигательных единиц обычно оказываются более низкопороговыми, т. е. вовлекаются в возбуждение раньше. Разница в возбудимости мотонейронов позволяет нервной системе дозировать силу сокращения, вовлекая в возбуждения меньшее или большее количество двигательных единиц мышцы. При длительных, но обычно слабых тонических напряжениях, связанных, например, с поддержанием позы, активируются только низко- пороговые медленные, устойчивые к утомлению двигательные единицы. Если необходимо осуществить сильное фазное напряжение, в возбуждение вовлекаются высокопороговые, быстрые сильные двигательные единицы.
Рассмотренные двигательные единицы теплокровных животных и человека относятся к классу так называемых фазных двигательных единиц. У амфибий и рептилий, а также в некоторых (немногих) мышцах теплокровных (наружные мышцы глаза) содержатся особые тонические двигательные единицы — мышечные волокна, которые существенно отличаются от волокон фазных единиц. Возбуждение тонических волокон не подчиняется закону «все или ничего» и имеет характер локального ответа, поэтому ограничивается областью нервно-мышечного окончания или тем участком, к которому непосредственно приложено электрическое или химическое раздражение. Охват возбуждением всего волокна возможен потому, что на каждом мышечном волокне имеется не одно, а множество нервных окончаний. Одновременное поступление к этим окончаниям нервного импульса вызывает сокращение всего волокна. Это сокращение существенно медленнее, чем сокращение фазных мышечных волокон.
Регистрация электрической активности двигательных единиц у человека показала, что в естественных условиях мышцы редко бывают полностью расслабленными. Обычно в них наблюдается небольшая, так называемая позная, активность, или позный тонус;при этом низкопороговые медленные двигательные единицы разряжаются с небольшой частотой.
Тонус и особенно его нарушения при ряде заболеваний нервной системы связаны с изменением состояния рефлекторных механизмов, в частности рефлексов с проприо- рецепторов мышц, повышение возбудимости которых ведет к повышению тонуса.
механизмы мышечного сокращения
Структура миофибрилл и ее изменения
при сокращении.Миофибриллы представляют собой сократительный аппарат мышечного волокна. В поперечнополосатых мышечных волокнах миофибриллы разделены на правильно чередующиеся участки (диски), обладающие разными оптическими свойствами. Одни участки анизотропны, т. е. обладают двойным лучепреломлением. В обыкновенном свете они выглядят темными, а в поляризованном — прозрачными в продольном направлении и непрозрачными в поперечном. Другие участки в обыкновенном свете выглядят светлыми — они изотропны,т. с. не обладают двойным лучепреломлением (рис. 34, а). Анизотропные участки обозначают буквой А,изотропные — буквой /. В середине диска Аразличается светлая полоска Н,
темная полоска Z, представляющая собой тонкую мембрану, сквозь поры которой проходят миофибриллы.
Благодаря наличию этой опорной структуры параллельно расположенные однозначные диски отдельных фибрилл внутри одного волокна во время сокращения не смещаются по отношению друг к другу. Современные представления о структуре миофибриллярного аппарата основываются на исследованиях структуры мышечного волокна при помощи электронной микроскопии, рентгеноструктурного анализа, фазово-контрастной и интерференционной микроскопии в сочетании с гистохимическими методами.
Установлено, что каждая миофибрилла мышечного волокна диаметром около 1 мкм состоит в среднем из 2500 протофибрилл,представляющих собой удлиненные полимери- зованные молекулы белков миозина и актина. Миозиновые протофибриллы, или, как их принято обозначать, нити, вдвое толще актиновых. Их диаметр примерно 10 нм. В состоянии покоя мышечного волокна нити расположены в миофибрилле таким образом, что тонкие длинные актиновые нити входят своими концами в промежутки между толстыми и более короткими миозиновыми нитями (рис. 34, б)'. Благодаря этому диски / состоят только из актиновых нитей, а диски А — из нитей миозина, а, возможно, еще и другого белка.
|
IIII1II1 | ||
|
|
■ |
|
|
|
в |
в§ |
|
| ||
|
Давая |
|
U-J |
Рис.34.
Структура миофибриллы (схема). Показаны
диски А и I полоски
7.и Н{а). Взаимное
расположение толстых {миозиновых) и ,
тонких
(актиновых) нитей в расслабленной
посередине диска / — (б)
и сокращенной (в) миофибрнлле.
Роль потенциала действия в возникновении мышечного сокращения.В естественных условиях деятельности скелетной мышцы инициатором ее сокращения является потенциал действия, распространяющийся при возбуждении вдоль поверхностной мембраны мышечного волокна.
Если кончик микроэлектрода при помощи микронанипулнтора приложить к поверхности мышечного волокна лягушки в области диска /, то при нанесении очень слабого электрического стимула, вызывающего деполяризацию, диски / по обе стороны от мембраны Z начнут укорачиваться. При этом, однако, сокращение распространяется не в стороны, а в глубь волокна, вдоль диска /. Приложение слабого стимула к другим участкам миофибриллы подобного эффекта не вызывает. Из этого следует, что деполяризация поверхностной мембраны мышечного волокна в области дисков / является пусковым механизмом сократительного процесса.
молекулами, устилающими внутреннюю поверхность его трубочек и цистерн. Концентрация свободных Са2+в полости ретикулума близка к концентрации их в наружной среде, т. е. во внеклеточной жидкости.
Важным
промежуточным звеном между деполяризацией
мембраны и началом мышечного
сокращения является проникновение в
область миофибрилл свободных ионов В
состоянии покоя основная часть ионов
Са2+в скелетном мышечном волокне
хранится в так называемом
саркоплазматическом ретикулуме.Он представляет собой замкнутую систему
внутриклеточных трубочек и цистерн,
окружающих каждую миофибриллу.
В
мембране саркоплазматического рётику-
лума локализованы две важнейшие
транспортные системы, обеспечивающие
накопление в ретикулуме ионов Са2,+(«секвестрация» — захват их из миоплазмы)
и освобождение Са2+из ретикулума
при возбуждении.
Функцию
кальциевого насоса выполняет так
называемая Са-зависимая АТФ-аза (Са —
АТФ-аза). Энергия, выделяющаяся при
расщеплении АТФ, используется для
секвестрации ионов
Са2+в ретикулум. Благодаря этому в покоящемся
волокне концентрация свободных ионов
Са2+в цитоплазме поддерживается
на очень низком уровне. Поступая внутрь
ретикулума (главным образом в его
продольные трубочки), ионы
Са2+частично связываются белковыми
1*11
с. 35. Схемагическое изображение вза-
иMoot ношений поверхностной
мембраны(1).
поперечных трубочек (2), боконых цистерн
13) и продольных трубочек (4)
саркоплазматического ретикулума
и миофибрилл (5) мышечного волокна.
дения потенциал действия с поверхностной мембраны распространяется вдоль мембраны поперечных трубочек в глубь волокна и при помощи особого, пока еще полностью не изученного, механизма вызывает освобождение ионов Са2+из боковых цистерн. Боковые цистерны расположены таким образом, что освободившиеся ионы Са2+попадают непосредственно в ту область, где происходит образование актомиозина.
Как отмечалось, начало мышечного сокращения приурочено к первой трети восходящего колена потенциала действия, а именно к моменту, когда внутренний потенциал волокна возрастает с исходных —90 мВ до примерно —50 мВ. Этот потенциал является пороговым для возникновения механического ответа. Предполагают, что именно при достижении указанного уровня деполяризации концентрация свободных ионов Са2+в миофибрилле достигает критической величины, необходимой для начала взаимодействия актиновых и миозиновых нитей.
Процесс освобождения Са2+прекращается после окончания пика потенциала действия. Тем не менее сокращение продолжает нарастать до тех пор, пока активация кальциевого насоса ретикулума не вызовет снижения концентрации ионов Са2+в миоплазме. Тогда сокращение сменяется расслаблением.
Таким образом, последовательность событий, ведущих к сокращению, а затем расслаблению мышечного волокна, представляется в следующем виде: раздражение —> возникновение потенциала действия —► проведение его вдоль клеточной мембраны и в глубь волокна по трубочкам—^освобождение Са2+из боковых цистерн саркоплазматического ретикулума и диффузия его к миофибриллам—► взаимодействие («скольжение») актиновых и миозиновых нитей, приводящее к укорочению миофибриллы —► активация кальциевого насоса—снижение концентрации свободных ионов Са2+в саркоплазме —► расслабление миофибрилл.
Роль АТФ в механизмах мышечного сокращения
В процессе взаимодействия миозиновых и актиновых нитей в присутствии Са2+ важную роль играет богатое энергией соединение — АТФ. Впервые В. А. Энгельгард и М. Н. Любимова в 1939 г. нашли, что мышечный белок — миозин — обладает свойствами фермента аденозинтрифосфатазы (АТФ-аза). Эти авторы также обнаружили, что под влиянием АТФ изменяются и механические свойства миозина — резко увеличивается растяжимость его нитей. В последующие годы был открыт белок актин и показано, что он активирует АТФ-азную активность миозина.
В настоящее время взаимодействие АТФ с сократительными белками и ионами Са2+ представляют следующим образом. В покоящейся мышце, т. е. в условиях, когда концентрация ионов Са2+в миоплазме очень низкая, взаимодействию миозиновых и актиновых нитей препятствуют молекулы белка тропонина, расположенные на актиновых нитях. Тропонин обладает очень высоким сродством к ионам Са2+. Как только концентрацияСа2+в миофибриллах повышается, тропонин связывает Са2+и изменяет свое расположение на актиновой нити таким' образом, что делает возможным ее взаимодействие с миози- новой нитью. Формирующиеся при этом поперечные мостики перемещают актиновую нить лишь на 1 % ее длины. Чтобы обеспечить дальнейшее продвижение нити и соответственно сокращение волокна, необходимо, чтобы эти мостики разъединились и прикрепились к новым участкам актиновой нити. Такое разъединение мостиков осуществляется при расщеплении молекул АТФ АТФ-азой миозина. Таким образом, взаимодействие миозина и актина, возможное при связывании Са2+тропонином, активирует АТФ-азу миозина, последняя расщепляет АТФ, а это приводит к разъединению миозина и актина. В присутствии Са2+и АТФ в миоплазме этот процесс многократно повторяется: мостики повторно образуются и расходятся, в результате чего актиновые нити «скользят» и мышечное волокно укорачивается.
Подсчитано, что при изотоническом сокращении скелетной мышцы лягушки поперечные мостики должны совершить за 0,1 с 50 таких движений, чтобы длина каждого саркомера волокна укоротилась на 50 %. При каждом движении мостиков происходит расщепление молекул АТФ.
Таким образом, присутствие АТФ в мышце является обязательным условием для обратимости связывания актина и миозина. Трупное окоченение возникает при условии, если концентрация АТФ в мышце падает ниже некоторой критической величины. Тогда комплекс актин — миозин становится стабильным.
Итак, энергия АТФ используется во время деятельности скелетной мышцы для трех основных процессов: 1) работы натр ий-калиевого насоса,обеспечивающего поддержание постоянства градиента концентрации ионовNa+ и К+по обе стороны мембраны; 2) процесса «скольжения» актиновых и миозиноеых нитей,ведущего к укорочению миофиб- рилл; 3) работы кальциевого насоса,необходимого для расслабления волокна. В соответствии с этим ферменты, расщепляющие АТФ, локализованы в трех различных структурах мышечного волокна: клеточной мембране, миозиновых нитях и мембранах саркоплаз- матического ретикулума.
Ресинтез АТФ, непрерывно расщепляющейся в процессе деятельности мышцы, осуществляется двумя основными путями. Первый состоит в ферментативном переносе фосфатной группы от богатого энергией фосфорного соединения креатинфосфатана аденозиндифосфорную кислоту. Креатинфосфат содержится в мышце в значительно больших количествах, чем АТФ, и обеспечивает ее ресинтез в течение тысячных долей секунды. Однако при интенсивной работе мышцы запасы креатинфосфата быстро истощаются, поэтому важен второй путь—более медленный ресинтез АТФ.Он связан с гликолитическими и окислительными процессами, протекающими в мышце как в условиях покоя, так и особенно интенсивно во время деятельности. Окисление молочнойи пиро- винограднойкислот, образующихся в мышце во время ее сокращения, сопровождается фосфорилированием аденозиндифосфорной кислоты и креатина, т. е. ресинтезом креатинфосфата и АТФ.
Нарушение ресинтеза АТФ ядами, подавляющими гликолитические и окислительные процессы, ведет к полному исчезновению АТФ и креатинфосфата, вследствие чего кальциевый насос перестает работать. Концентрация Са +в области миофибрилл значительно возрастает и мышца приходит в состояние длительного необратимого укорочения.
Теплообразование при сократительном процессе и
энергия сокращения
Образование тепла в мышечной ткани при работе было открыто Гельмгольцем и В. Я. Данилевским во второй половине XIX в. В дальнейшем Хиллу с сотр. удалось создать высокочувствительные приборы, которые позволили зарегистрировать и измерить теплопродукцию мышц и нервов в покое и при возбуждении.
По своему происхождению и времени развития теплообразование было подразделено Хиллом на две основные фазы. Первая фазаприблизительно в 1000 раз короче второй и называется фазой начального теплообразования. Она начинается с момента возбуждения мышцы и продолжается в течение всего сокращения, включая фазу расслабления.
В свою очередь начальное теплообразование может быть разделено на несколько частей: а) тепло активации; б) тепло укорочения; в) тепло расслабления.
Тепло активации освобождается непосредственно после нанесения раздражения, но до сколько-нибудь различимого сокращения мышечных волокон. Поэтому указанная порция тепла рассматривается как тепловой эффект тех химических процессов, которые переводят мышцу из невозбужденного состояния в активное. При тетаническом сокращении тепло активации выделяется в течение всего времени раздражения мышцы при каждом потенциале действия.
Тепло укорочения обусловлено самим сократительным процессом. Если путем сильного растяжения мышцы воспрепятствовать ее сокращению, эта порция тепла не выделяется.
Тепло расслабления связано с освобождением энергии в результате расслабления мышцы. Если мышца подняла груз во время сокращения, то по окончании его количество выделяемого тепла увеличивается.
Вторая фазатеплопродукции длится несколько минут после расслабления и носит название запаздывающего, или восстановительного, теплообразования.
Она связана с химическими процессами, обеспечивающими ресинтез АТФ. В опытах на мышцах, сокращающихся в отсутствие кислорода, Хилл показал, что в отличие от начального теплообразования, для которого кислород не нужен, 90 % восстановительного тепла образуется в результате окислительных процессов и лишь 10 % этого тепла обусловлены анаэробными процессами обмена веществ. Тепло восстановления по своей величине примерно равно количеству тепла, выделяемого мышцей во время сокращения. Это соответствие становится понятным, если учесть, что химические процессы, обусловливающие восстановительное теплообразование, направлены на ресинтез АТФ, являющийся основным непосредственным источником энергии мышечного сокращения. Главную роль в ресинтезе АТФ и восстановительном теплообразовании играют процессы гликолиза и окислительного фосфорилирования. Отравление мышцы монойодуксусной кислотой, прекращающей гликолитическое образование молочной и пировиноградной кислот, почти полностью выключает запаздывающее теплообразование и ресинтез АТФ даже в присутствии кислорода.
РАБОТА И СИЛА МЫШЦ
Величина сокращения (степень укорочения) мышцы при данной силе раздражения (т. е. при данном числе активированных волокон) зависит как от ее морфологических свойств, так и от физиологического состояния. Длинные мышцы сокращаются на большую величину, чем короткие. Умеренное растяжение мышцы увеличивает ее сократительный эффект; при сильном растяжении сокращение мышцы ослабляется. Если в результате длительной работы развивается утомление мышцы, то величина ее сокращения снижается.
Для измерения силы мышцы определяют либо тот максимальный груз, который она в состоянии поднять, либо максимальное напряжение, которое она может развить в условиях изометрического сокращения. Эта сила может быть очень велика. Установлено, что собака мышцами челюсти может поднять груз, превышающий массу ее тела в 8,3 раза.
Одиночное мышечное волокно способно развить напряжение 100—200 мг. Учитывая, что общее количество мышечных волокон в теле человека приблизительно 15—30 млн., они могли бы развить напряжение в 20—30 т, если бы все одновременно тянули в одну сторону.
Сила мышцы при прочих равных условиях зависит от ее поперечного сечения. Чем больше физиологическое поперечное сечение мышцы, т. е. сумма поперечных сечений всех ее волокон, тем больше тот груз, который она в состоянии поднять. Физиологическое поперечное сечение совпадает с геометрическим только в мышцах с продольно расположенными волокнами; у мышц с косым расположением волокон сумма поперечных сечений может значительно превышать геометрическое поперечное сечение самой мышцы (рис. 36). По этой причине сила мышцы с косо расположенными волокнами значительно больше, чем сила мышцы той же толщины, но при продольном расположении волокон. Чтобы иметь возможность сравнивать силу разных мышц, максимальный груз, который в состоянии поднять мышца, делят на площадь ее физиологического поперечного сечения. Таким образом вычисляют абсолютную мышечную силу. Абсолютная сила, выраженная в килограммах на I см2, икроножной мышцы человека равна 5,9, сгибателя плеча — 8,1, жевательной мышцы — 10, двуглавой мышцы плеча — 11,4, трехглавой мышцы плеча — 16,8, гладких мышц— 1.
Примером мышц с продольным расположением волокон может служить m.sartorius, с косым —m.m.intercostales. Большинство мышц млекопитающих и человека перистого
строения. Перистая мышца имеет большое физиологическое сечение, а потому обладает большой силой.
Зависимость
а
— мышцы с параллельным ходом волокон,
6 — веретенообразная мышца; в -- перистая
мышца.
Если покоящуюся мышцу постепенно растягивать за сухожильные концы, то в силу эластических свойств напряжение мышцы будет возрастать так, как это показано на рис. 37 (кривая А).
Нанесение электрических раздражений на мышцу вызывает прирост напряжения (кривая Б), величина которого с увеличением исходной длины мышечных волокон вначале возрастает, а затем уменьшается. Если сопоставить кривую Б с кривой А, то можно получить представление о зависимости этого активного прироста изометрического напряжения от длины мышечных саркомеров. При некоторой средней длине саркомеров сила, развиваемая мышечным волокном, максимальна. При перерастяжении волокон эта сила уменьшается. Причина такой зависимости напряжения от исходной длины саркомеров становится ясной при рассмотрении взаимного расположения миозиновых и актиновых нитей при разной длине саркомеров (рис. 37,В).
О
Рис. 37. Взаимоотношения между силой и длиной мышцы.
Рис.
36. Типы строения различных мышц (гю А.
А. Ухтомскому).

