

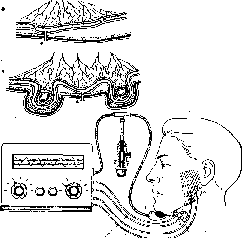
Рис.
174. Регистрация функции жевательной
мускулатуры методом миоэлектромастмкациа-
графин (схема).
□
□ □ □□□□□□□
о^а
□
х^'Ъ
□^□□□□□□□□□□□□□cjI датчик мастика циографа; 2 — электроды лля отведении биопотенциалов жеигмельиых мышц.
/ i
~ ,

Рис.
176. Зоил длн определения кислотности
разных .чон желудка (по Линару).
I — штепсель л л и сиединения электродов с регистратором рН; 2 — решило ил я i рубка с отверстиями (3), соединяющая резиновый баллон(G) с регистра тиром; 4 релиновый зонд; 5 — провода; 7—8 — рИ-олнцм: 9 — сурьмяные электроды; 10 - - кялпмелепые электроды.
Изучение деятельности пищеварительной системы человека требует с п е- циальныхметодическихиодхолов.
Акт жевания исследуют путем регистрации движений нижней челюсти (мастикаци- ография)иногда с одновременной электромиографией жевательных мышц (рис. 174). При помощи капсул Лейиш — Красногорского(рис. 175) можно собрать раздельно слюну околоушной, подчелюстной и подъязычной желез.
Изучение пищеварительных органов, расположенных в брюшной полости, требует иных методических приемов. Для учета секреторной деятельностипищеварительных желез человека используются зондовые и беззондовые методы. В методах первой группы обследуемый проглатывает резиновую трубку (или вводят ее через нос), один конец которой достигает полости желудка или двенадцатиперстной кишки (зонд может быть проведен и ниже). При помощи специального зонда определяют рН в желудке и верхних отделах кишечника (рис. 176). Зонд может иметь несколько датчиков рН, расположенных на различных уровнях. Получаемые данные характеризуют кислото- выделительную деятельность, эвакуацию содержимого желудка и кишечник, транзит кислой жидкости через двенадцатиперстную кишку и т.д.
Рис.
175. Собирание слюны у человека с помощью
капсулы. Л с шли Красногорского.
Вил капсулы изображен в нижней части рисунка. I трубка для отсасывании аоз- духа из внешней камеры капсулы; 2 — трубка для оттока слюны из внутренней камеры капсулы.
С
развитием радиоэлектроники получила
применение радиотелеметрическая
методика. Сущность ее заключается в
том, что человеку дают проглотить
миниатюрный радиопередатчик —
радиопилюлю.Она состоит
из генератора электромагнитных
колебаний, источника питания (сухой
элемент или аккумулятор) и датчика. Под
влиянием воспринимаемых параметров
датчик меняет частоту излучаемых
радиопилюлей колеба-
Рис.
177. Электрогастрография.
а
— активный электрод на коже в области
желудка; б. в - индифферентные электроды
на нижних конечностях (по М. В.
Собакину).Блок-схема элеитрогастрографа эгс-3

Электрогастрографа
ний. Они воспринимаются антенной, надетой на обследуемого, и радиоприемником с записывающим устройством. Радиопилюля свободно проходит по желудочно-кишечному тракту. С ее помощью можно оценить секреторную деятельность желудка и моторную активность его и кишечника, а также гидролиз ряда питательных веществ.
Моторную активность желудка можно изучать электрогастрографически,отводя с кожи живота и конечностей человека биопотенциалы (рис. 177), генерируемые гладкими мышцами желудка. Этот метод модернизируется для регистрации моторной активности тонкой, толстой кишки и желчного пузыря.
Для изучения моторной активности органов пищеварения человека широкое применение нашли рентгенологические методы (рентгеноскопия, рентгенография, рентгено- кинематогр афия).
Следует отметить также методы эндоскопии,которые позволяют осмотреть, сфотографировать слизистую оболочку желудка и начального отдела кишечника, устья выводных протоков, взять для исследования маленький кусочек слизистой оболочки (биопсия). Взятые кусочки подвергают гистологическому и биохимическому исследованию.
Для характеристики гидролиза белков и всасывания аминокислот широкое распространение получили пробы с мечеными белками (казеином или альбумином).
Гидролиз и всасывание жиров исследуют также методами с дачей обследуемым меченых и немеченых жиров с последующим динамическим учетом содержания хило- микронов (микрокапельки жира, окруженные липопротеидной мембраной) или общих липидов в сыворотке крови. Широко применяется учет жира в кале.
Существует и ряд других методов, подробно описанных в специальных курсах.
ПИЩЕВАРЕНИЕ В ПОЛОСТИ РТА
Переработка принятой пищи начинается в полости рта. Здесь происходят ее измельчение, смачивание слюной, анализвкусовых свойств пищи, начальный гидролизнекоторых пищевых веществ и формирование пищевого комка.Средняя длительность пребывания пищи в полости рта 15—18 с.
I
II III IV V
Рис.
178. Кимограмма жевательного периода
(по Рубинову).
Фазы:
I _ покоя:
П — введения пиши в рот; 1П — ориентировочная;
IV — основная: V - формирования пищевого
комка и проглатывания пиши, внизу -
отметка времени I с.
Поступившая в рот пища раздражает вкусовые, тактильные и температурные рецепторы. Вкусовые рецепторы расположены преимущественно в сосочках языка и рассеяны в слизистой оболочке полости рта, тактильные, температурные и болевые — по всей слизистой оболочке. Сигналы от этих рецепторов по центростремительным нервным волокнам тройничного, лицевого и языкоглоточного нервов доходят до нервных центров ряда рефлексов. Центробежные импульсы от этих центров рефлекторно возбуждают секрецию слюнных, желудочных и поджелудочной желез, выход желчи в двенадцатиперстную кишку, изменяют моторную активность желудка. Раздражение рецепторов полости рта имеет важное значение в осуществлении актов жевания и глотания. Таким образом, несмотря на то что пребывание пищи во рту кратковременно, этот отдел пищеварительного тракта оказывает влияние на все этапы переработки пищи.
Жевание.В полость рта пища поступает в виде кусков, смесей разного состава и консистенции или жидкостей. В зависимости от этого пища или сразу проглатывается, или подвергается предварительной механической и химической обработке.
Акт жевания совершается рефлекторно. Находящаяся во рту пища раздражает рецепторы, от них сигналы по афферентным волокнам тройничного нерва передаются в центр жевания, а от него по эфферентным волокнам тройничного нерва — к жевательным мышцам. В координации акта жевания имеют также важное значение сигналы от проприорецепторов жевательных мышц.
Жевательный период имеет фазы различной длительности в зависимости от свойств пережевываемой пищи (рис. 178) (покой, введение пищи в рот, ориентировочная, основная, формирование пищевого комка и глотание).
Слюноотделение.На начальном этапе пищеварения велика роль слюны. Она продуцируется тремя парами крупных слюнных желез: околоушными, подчелюстными и подъязычными — и множеством мелких железок, находящихся на поверхности языка, в слизистой оболочке неба и щек. Из желез по выводным протокам слюна поступает в полость рта. В зависимости от вырабатываемого секрета слюнные железы бывают трех типов: серозные(вырабатывают жидкий секрет, не содержащий слизи — муцина);смешанные(вырабатывают серозно-слизистый секрет) и слизистые(вырабатывают слюну, богатую муцином). Околоушная железа и малые железы боковых поверхностей языка имеют серозные клетки и продуцируют жидкую слюну. Слизистые железы расположены на корне языка и неба. В подчелюстной и подъязычной железах имеются серозные и Слизистые клетки, поэтому их называют смешанными. Смешанные железы находятся также в слизистой оболочке кончика языка, щек, губ. Из ацинусов слюнных желез секрет поступает в систему укрупняющихся протоков, собирающихся в общий выводной проток, выносящий слюну в полость рта. Вне приема пищи у человека слюна выделяется в среднем 0,24 мл/мин для увлажнения полости рта, при жевании — 3—3,5 мл/мин (около 200 мл/ч) в зависимости от вида принимаемой пищи. В ответ на вводимый раствор лимонной кислоты слюноотделение может достигать 7,4 мл/мин. За сутки продуцируется 0,5—2,0 л слюны, около трети ее образуется околоушными железами.
Состав и свойства слюны.Слюна представляет собой вязкую слегка опалесцирую- гцую и мутноватую жидкость с плотностью 1,001 —1,017. Состав слюны в большей мере зависит от скорости ее секреции; рН смешанной слюны 5,8—7,4; рН слюны околоушных желез ниже (5,81), чем подчелюстных (6,39). С увеличением скорости секреции рН повышается до 7,8.
Смешанная слюна содержит 99,4—99,5% воды, остальное — сухой остаток. Неорганические компоненты слюны: хлориды и карбонаты, фосфаты и другие соли натрия, калия, кальция, магния и др. Концентрация электролитов неодинакова в слюне разных слюнных желез и увеличивается с повышением скорости ее секреции, но слюна и в этих условиях имеет более низкое осмотическое давление, чем плазма крови.
Слюна содержит органические вещества, которых в 2—3 раза больше, чем минеральных солей. Органические вещества являются продуктом секреторной деятельности слюнных желез, обмена веществ в них и частично транспортируются из крови. В составе слюны выделяются различные белки, свободные аминокислоты, некоторые углеводы, мочевина, аммиак, креатинин и другие вещества. Слюна содержит муцин, который придает ей вязкость, благодаря наличию муцина пропитанный слюной пищевой комок легко проглатывается.
Слюна достаточно богата ферментами, хотя содержание некоторых из них невелико. Слюна человека обладает способностью активно гидролизовать углеводы. Это осуществляется а-амилазой,расщепляющей полисахариды (крахмал, гликоген) с образованием декстринов, а затем дисахаров (мальтозы) и частично глюкозы. Дисахаридазная активность слюны низкая.
Амилаза слюны начинает свое действие в полости рта, но оно незначительно вследствие кратковременного пребывания здесь пищи. Гидролиз углеводов ферментами слюны продолжается в желудке, пока в глубокие слои его пищевого содержимого не проникает кислый желудочный сок, прекращающий действие карбогидраз и инактивирующий их.
В слюне содержится ряд других ферментов: протеиназы(катепсины, саливаин, глан- дулаин), липазы,щелочная и кислая фосфатазы, РНК-азы.Не исключено, что и они принимают участие в процессе пищеварения, но активность их невелика. Слюна обладает бактерицидным свойством за счет содержащегося в ней фермента лизоцима (мурамидаза). В слюне содержится калликреин,который принимает участие в образовании кининов, расширяющих кровеносные сосуды, что может иметь значение в увеличении кровоснабжения слюнных (и других) желез при приеме пищи.
Ферментный состав и свойства слюны изменяются с возрастом человека, зависят от режима питания и вида пищи. На пищевые вещества выделяется более вязкая слюна и тем больше, чем суше принимаемая пища. На отвергаемые вещества, кислоты, горечи
выделяется значительное количество более жидкой слюны. Адаптация слюноотделения к виду принимаемой пищи выражается не только в изменении объема и вязкости слюны, но и ее каталитической активности. Количество и состав слюны в связи с приемом пищи определяются регуляторными воздействиями на слюнные железы.
Регуляция слюноотделения
Прием пищи возбуждает слюноотделение рефлекторно.При достаточно сильном раздражении и высокой возбудимости пищевого центра слюноотделение начинается через 1—3 с, а при слабой силе раздражения латентный период достигает 20—30 с. Слюноотделение продолжается весь период еды и прекращается вскоре после нее.
От рецепторов полости рта сигналы передаются в ЦНС по афферентным волокнам тройничного, лицевого, языкоглоточного и блуждающего нервов. Основной слюноотделительный центр расположен в продолговатом мозге. Именно сюда, а также в боковые рога верхних грудных сегментов спинного мозга поступают сигналы из полости рта и расположенных выше отделов мозга. Отсюда влияния по эфферентным парасимпатическим и симпатическим нервным волокнам направляются к слюнным железам.
Парасимпатическая иннервация слюнных желез начинается из ядер продолговатого мозга (рис. 179). Симпатическая иннервация слюнных желез осуществляется от боковых рогов II—IV грудных сегментов спинного' мозга.
Рис.
179. Схема путей рефлекторного возбуждения
подчелюстной (а) и околоушной (й) слюнных
желез.
а: I — тройничный нерв; 2 — узел тройничного нерва (Гассеров); 3 — ядро лицевого нераа; 4 - - лицевой нерв; 5 — коленчатый узел: Ь барабанная струна: 7 - язычный пери; 8 - подчелюстная железа и подчелюстной ганглий; 9 — крыло небный ганглий. 10 — центр слюноотделения, б: 1 — гройпичнмй > »ел; 2 тройничный нерв; 3 -- ядро языкоглоточного нерва; 4 — лицевой нерв; 5 — коленчатый узел; 6 - верхнечелюстной нерв: 7 — ушной узел; 8 — ушио-височный нерв; 9 - барабаннаи струпа; Ю - язычный нерв; 11 — пещеристый узел; 12 - нзыкоглоточный нерв, 13 - околоушная слюнная железа. Выделены чу нет пи дельные нервные волокна, идущиеor рецепторных аппаратов я^мка, и преганглионарные парасимпатические волокна; пунктирная линия — постганглиопарныс парасимпатические волокна.
В окончаниях постганглионарных парасимпатических волокон высвобождается медиатор — ацетилхолин, возбуждающий секреторные клетки слюнных желез. При этом выделяется большое количество жидкой слюны. Длительное раздражение парасимпатических нервов вызывает истощение слюнных желез: снижается содержание в слюне органических компонентов, в гландулоцитах резко уменьшается количество гранул.
В окончаниях симпатических нервных волокон высвобождается норадреналин. Под его влиянием выделяется небольшое количество густой слюны. Раздражение симпатических нервов усиливает образование гранул секрета в гландулоцитах. Одновременное или предшествующее раздражение парасимпатических нервов усиливает симпатические секреторные эффекты. Ответная реакция при раздражении симпатических нервов возникает спустя более длительный латентный период, чем после раздражения парасимпатических. •

Слюноотделение начинается по типу условных рефлексов— в ответ на вид и запах пищи.
Рефлекторные влияния могут и тормозить слюноотделение, вплоть до его прекращения. Такое торможение может быть вызвано болевым раздражением, отрицательными эмоциями, умственным напряжением, дегидратацией организма. Все эти воздействия снижают активность пищевого центра и его части — центра слюноотделения. Возбудителями последнего могут быть некоторые гуморальные вещества. Так, обильное отделение слюны наблюдается при асфиксии вследствие раздражения центра слюноотделения угольной кислотой. Раздражают его некоторые токсины и при этом наблюдается обильное слюноотделение.
Перерезка парасимпатических нервов, иннервирующих слюнные железы, временно резко усиливает выделение ими слюны — паралитическая секреция. В первые три дня непрерывное слюноотделение обусловлено повышенным выделением ацетилхолина вследствие дегенерации нейронов, поэтому эта секреция называется дегенеративной. В последующие дни паралитическая секреция связана с повышенной чувствительностью денервированной железы к ряду веществ, циркулирующих в крови, к которым железа с нормальной иннервацией малочувствительна. Холиномиметические фармакологические вещества вызывают усиление слюноотделения, холинолитические — тормозят его.
Глотание
Глотание является рефлекторным актом, центр его находится в продолговатом мозге, на дне IV желудочка. Доказательством рефлекторной природы глотания служит следующее: если смазать корень языка и глотки раствором кокаина и выключить таким образом рецепторы их слизистой оболочки, то глотание осуществляться не будет. Глотание невозможно и после перерезки эфферентных нервов глотки.
Рефлекс глотания состоит из ряда последовательных звеньев. Строгая координация звеньев глотательного рефлекса обеспечивается сложными взаимосвязями различных отделов ЦНС — от продолговатого мозга до коры большого мозга. Центр глотания связан с центром дыхания, что обеспечивает задержку дыхания во время глотания и имеет значение в предотвращении попадания пищи в воздухоносные пути. Связь центра глотания с центром регуляции сердечной деятельности подтверждается учащением сердечных сокращений во время глотания.
Акт глотания делится на три фазы: 1) ротовую(произвольную), 2) глоточную(быструю непроизвольную), 3) пищеводную(медленную непроизвольную). Акт глотания изучен рентгеновским методом (рис. 180).
Из пигцевой пережеванной массы во рту отделяется пигцевой комок объемом 5—15 см3,который движениями языка и щек перемещается на спинку языка. Сокращениями передней части языка пищевой комок прижимается к твердому небу, затем последовательными сокращениями средней части отжимается кзади и переводится на корень языка за передние дужки (1-я фаза).
Раздражение рецепторов слизистой оболочки корня языка рефлекторно вызывает сокращение мыщц, приподнимающих мягкое небо, и мышц языка, что препятствует попаданию пищи в полость носа. Движения языка способствуют проталкиванию пиши в полость глотки. Одновременно с этим происходит сокращение мышц, смещающих подъязычную кость и вызывающих поднятие гортани, вследствие чего закрывается вход в дыхательные пути, что препятствует поступлению в них пищи. Переводу пищи в глотку способствует повышение давления в полости рта. Возвращению пищи из глотки в полость рта препятствуют поднявшийся вверх корень языка и плотно прилегающие к нему дужки. Как только пища поступила в полость глотки, начинают сокращаться мышцы, суживающие просвет глотки выше пищевого комка, вследствие чего последний передвигается в пищевод. Существенное значение при этом имеет градиент давления между полостью глотки и началом пищевода. Перед глотанием глоточно-пищеводный сфинктер закрыт, во время глотания давление в глотке повышается до 6 Па (45 мм рт. ст.) и через открывшийся сфинктер пищевой комок поступает в пищевод (2-я фаза), где давление в это время не более 4 Па (30 мм рт. ст.). Давление в глотке значительно выше, сфинктер закрыт и невозможно обратное забрасывание пищевого комка в глотку. Две фазы глотательного цикла длятся около 1 с.
ис. 180. Последовательность движений в полости
Вторую фазу глотания нельзя выполнить произвольно, если в полости рта нет пищи или слюны. В этом легко убедиться, делая подряд несколько глотательных движений: первое движение происходит легко, так как в полости рта всегда имеется небольшое количество слюны; при отсутствии слюны, которая проглочена, дальнейшие глотательные движения осуществить не удается.
Третью фазу глотания составляют прохождение пищи по пищеводу и перевод ее в желудок. Движения пищевода взаимосвязаны с движениями глотательного аппарата и вызываются рефлек- торно при каждом глотательном акте. Если в эксперименте на собаках перерезать пищевод и вкладывать пищу непосредственно в него, то она передвигается .в сторону желудка только после совершения глотательных движений. Продолжительность продвижения по пищеводу твердой пищи в среднем 8—9 с, жидкой 1—2 с.
В момент акта глотания пищевод подтягивается к зеву и начальная часть его расширяется, принимая пищевой комок.
Сокращения пищевода имеют характер волны, возникающей в верхней его части и распространяющейся в сторону желудка. При этом последовательно сокращаются кольцеобразно расположенные мышцы пищевода (в верхней трети поперечнополосатые, в нижних двух третях — гладкие), передвигая перед волной сокращения находящийся в несколько расширенной части пищевода пищевой комок в сторону желудка. Такой тип сокращений называется перистальтическим.
Первичная перистальтическая волна, вызываемая актом глотания, доходит до уровня пересечения пищевода с дугой аорты. Далее возникает вторичная перистальтическая волна, вызываемая не актом глотания, а первичной перистальтической волной. Вторичная волна продвигает пищевой комок до кардиальной части желудка. Средняя скорость распространения перистальтической волны по пищеводу человека 2—5 см/с, она в большой мере зависит от свойств пищи. Вторичная перистальтическая волна может быть вызвана остатком пищевого комка в нижней трети пищевода, благодаря чему этот остаток переводится в желудок.
Регуляция моторной деятельности пищевода осуществляется в основном эфферентными волокнами блуждающего и симпатического нервов; определенная роль принадлежит интрамуральным нервным образованиям пищевода.
Вне глотательных движений вход из пищевода в желудок закрыт. Когда пища и перистальтическая волна достигают конечной части пищевода, происходит рефлекторное понижение тонуса мышц кардии и пищевой комок переходит в желудок. При наполнении желудка тонус кардии повышается, что предупреждает забрасывание содержимого желудка в пищевод. Парасимпатические волокна блуждающего нерва стимулируют перистальтику пищевода и расслабляют кардию, симпатические волокна тормозят моторику пищевода, но повышают тонус кардиальной части.
ПИЩЕВАРЕНИЕ В ЖЕЛУДКЕ
Желудок принимает участие в пищеварительном конвейере, а также в межуточном обмене веществ, поддержании постоянства рН крови и кроветворении (вырабатывает желудочный мукопротеид, или внутренний фактор Касла). Пищеварительными функциями желудка являются: депонирование пищи,ее механическая и химическая обработка,постепенная порционная эвакуацияпищевого содержимого в кишечник. Находясь в желудке в течение нескольких часов, пища набухает, разжижается, многие ее компоненты растворяются и подвергаются гидролизу ферментами слюны и желудочного сока. Желудочный сок обладает также антибактериальным действием.
Карбогидразы слюны продолжают действовать на углеводы пищи, находящиеся в центральной части пищевого содержимого желудка (куда еще не диффундировал кислый желудочный сок, прекращающий действие карбогидраз слюны). Ферменты кислого желудочного сока воздействуют на пищевые белки в относительно узкой зоне пищевого содержимого, находящегося в Непосредственном контакте со слизистой оболочкой желудка и в небольшом удалении от нее, куда диффундировал желудочный сок и не был нейтрализован за счет буферных свойств пищи. Ширина этой зоны зависит от количества и свойств желудочного сока и принятой пищи. Вся масса пищи в желудке не смешивается с соком. По мере разжижения и химической обработки пищи ее слой, прилегающий к слизистой оболочке, движениями желудка перемещается в его антральную часть, откуда эвакуируется в кишечник. Таким образом, пищеварение в желудке (по типу полостного) осуществляется некоторое время за счет слюны, но ведущее значение имеют секреторная и моторная деятельность самого желудка.
Секреторная деятельность желудка.
Состав и свойства желудочного сока
Желудочный сок продуцируется железами желудка, расположенными в слизистой оболочке. В области свода желудка железы имеют в своем составе главные гландулоциты (главные клетки),продуцирующие пепсиногены; париетальные гландулоциты (обкла- дочные клетки),которые синтезируют и выделяют соляную кислоту, и мукоциты (добавочные клетки),выделяющие мукоидный секрет. В пилорической части желудка париетальных гландулоцитов нет. В желудок мукоидный секрет и электролиты выделяются также клетками поверхностного эпителия. Вследствие различия в строении фундальных и пило- рических желез они продуцируют сок разного состава. Ведущее значение в желудочном пищеварении имеет фундальный желудочный сок.
В желудке человека выделяется 2,0—2,5 л желудочного сока в сутки. Он представляет собой бесцветную прозрачную жидкость, содержащую соляную кислоту (0,3—0,5%), и потому имеет кислую реакцию (рН 1,5—1,8). рН пищевого содержимого желудка значительно выше, так как сок фундальных желез частично нейтрализуется принятой пищей.
Считают, что соляная кислота продуцируется постоянной концентрации, но кислотность желудочного сока широко варьирует в результате изменения числа одновременно функционирующих париетальных гландулоцитов и нейтрализации соляной кислоты щелочными компонентами. Чем быстрее выделяется желудочный сок, тем меньше он нейтрализуется и тем выше его кислотность.
Для характеристики кислотности желудочного сока определяют: концентрацию ионов водорода, концентрацию неионизированного водорода и титруемую кислотность, которая является суммой концентрации водородных ионов и недиссоцГшрованной кислоты. С учетом объема секреции кислотовыделение оценивается дебитом свободной и связанной соляной кислоты.
Соляная кислотажелудочного сока вызывает денатурацию и набухание белкови тем самым способствует их последующему расщеплению пепсинами, активирует пепсиногены,создает кислую среду, необходимую для расщепления пищевых белков пепсинами; участвует в антибактериальном действиижелудочного сока и регуляции деятельности пищеварительного тракта(в зависимости от величины рН пищевого содержимого нервные механизмы и гастроинтестинальные гормоны усиливают или тормозят деятельность желудка).
Кислотность желудочного сока ребенка первых месяцев жизни низкая, она возрастает к концу года и к 7—12 годам достигает показателей, характерных для взрослых.
В желудочном соке имеются многие неорганические вещества: хлориды, сульфаты, фосфаты, бикарбонаты натрия, калия, кальция и магния, аммиак. Осмотическое давление желудочного сока выше, чем плазмы крови.
Органические компоненты желудочного сока представлены большим числом азотсодержащих веществ (200—500 мг/л): мочевиной, мочевой и молочной кислотами, аминокислотами, полипептидами. Содержание белков достигает 3 г/л, мукопротеидов — до 0,8 г/л, мукопротеаз — до 7 г/л. Органические вещества являются продуктами секреторной деятельности желудочных желез и обмена веществ в слизистой оболочке желудка, а также транспортируются через нее из крови. Особое значение для пищеварения имеют ферменты.
Главные гландулоциты желудочных желез человека синтезируют и выделяют пепсиногены двух групп.Пепсиногены первой группы (их 5) образуются в своде желудка, второй группы (их 2) — в привратниковой (пилорической) части желудка и начальной части двенадцатиперстной кишки. При активации пепсиногенов в кислой среде путем отщепления от них полипептида, являющегося ингибитором пепсина, образуется несколько пепсинов. Собственно пепсинамипринято называть ферменты, гидролизующие белки с максимальной скоростью при рН 1,5—2,0. Другая их фракция гидролизует белки при оптимальном рН 3,2—3,5 и называется гастриксином.Отношение между пепсином и гастри- ксином в желудочном соке человека от 1:2 до 1:5. Эти ферменты различаются действием на разные виды белков. Пепсины обладают выраженным свойством створаживать молоко. Возможность действия пепсинов в широком диапазоне рН имеет большое значение в желудочном протеолизе, происходящем при разном рН в зависимости от объема и кислотности желудочного сока, буферных свойств и количества принятой пищи. Протеазы желудочного сока расщепляют белки до крупных полипептидов (аминокислот при этом освобождается мало). Однако белки, подвергнутые предварительному действию желудочных протеаз и образовавшиеся при этом «осколки» белковой молекулы, затем легче расщепляются протеазами сока поджелудочной железы и тонкой кишки.
Желудочный сок взрослого человека обладает небольшой липолитической активностью.Эта липолитическая активность имеет важное значение для ребенка в период его молочного вскармливания (расщепление уже эмульгированных жиров молока).
Железы, расположенные в области малой кривизны желудка, продуцируют секрет с более высокой кислотностью и содержанием пепсина, чем железы большой кривизны желудка.
Важным компонентом желудочного сока являются мукоиды.Слизь, содержащая мукоиды, защищает оболочку желудка от механических и химических раздражений. Секреция слизи стимулируется местным раздражением слизистой оболочки, удалением слизи с ее поверхности, блуждающими и чревными нервами. К числу мукоидов относится и гастромукопротеид (внутренний фактор Касла).
Железы пилорической части желудка выделяют небольшое количество сока слабощелочной реакции с большим содержанием слизи. Секрет пилорических желез обладает небольшой протеолитической, липолитической и амилолитической активностью. Часть ферментов синтезируется непосредственно в клетках пилорических желез, а часть выделяется ими из крови. Существенного значения в желудочном пищеварении эти ферменты не имеют.
Щелочной пилорический секрет частично нейтрализует кислое содержимое желудка, эвакуируемое из желудка в двенадцатиперстную кишку.
Регуляция желудочной секреции
Вне пищеварения железы желудка человека выделяют небольшое количество желудочного сока. Прием пищи резко увеличивает его выделение железами тела желудка (но не пилорическими) в результате стимуляции желудочных желез нервными и гуморальными механизмами, составляющими единую систему регуляции. Стимулирующие и тормозные регуляторньге факторы обеспечивают зависимость сокоотделения желудка от вида принимаемой пищи. Указанная зависимость была впервые обнаружена в лаборатории И. П. Павлова в опытах на собаках с изолированным павловским желудочком (рис. 181). Не только объем и длительность секреции, но и кислотность, и содержание в соке пепсинов определяются характером принятой пищи. Так, по данным павловской лаборатории, показатели секреции на три пищевые раздражителя (мясо, хлеб, молоко) располагались следующим образом в порядке убывания:
Длительность секоении Кислотность сока Переваривающая сила сока
А/ГСТРП ттрГ5 Л/Гптттгп
Хлеб Мясо Молоко
Мясо Молоко Хлеб
Хлеб Мясо Молоко
На все виды раздражителей выделяется пепсина больше в начале секреции и меньше при ее завершении. Пищевые раздражители, вызывающие секрецию с большим участием в ней блуждающих нервов (хлеб), стимулируют выделение сока с более высоким содер-