

Вірусологія
.pdfКількісний вміст кожного з компонентів в різних сумішах середовищ значно варіює. Цей тип середовищ використовують рідко.
Насьогодні, в основному, використовують штучні поживні середовища. До напівсинтетичних поживних середовищ відносять гідролізати різних білкових продуктів (гідролізат лактальбуміну, гемогідролізат, м’язевий ферментативний гідролізат, амінопептид та ін.). Серед синтетичних поживних середовищ найбільш широко застосовують середовище 199 (табл. 2.14.) та середовище Ігла. До їх складу входить більш ніж 60 компонентів (амінокислоти, вітаміни, компоненти нуклеїнових кислот, джерело ліпідів та вуглеводів, мінеральні солі та ін.).
|
|
|
Таблиця 2.14 |
||
Середовище 199 (Морган, Мортон, Паркер; 1950 р.) |
|||||
|
|
|
|
|
|
Амінокислоти |
мг |
Джерела ліпідів |
|
мг |
|
L–Аргінін- |
70 |
Твін-80 |
|
20 |
|
монодихлорид |
|
|
|
|
|
L-Гістидин- |
20 |
Холестерин |
|
0,2 |
|
монодихлорид |
|
|
|
|
|
L-Лізин- |
70 |
Компоненти |
|
мг |
|
монондихлорид |
|
нуклеїнових кислот |
|
|
|
DL – Триптофан |
20 |
Аденін сульфат |
|
10 |
|
DL – Фенілаланін |
50 |
Ксантин |
|
0,3 |
|
DL – Метіонін |
30 |
Гіпоксантин |
|
0,3 |
|
DL – Серин |
50 |
Тимін |
|
0,3 |
|
DL – Треонін |
60 |
Урацил |
|
0,3 |
|
DL – Лейцин |
120 |
Гуанін-гідрохлорид |
|
0,3 |
|
DL – Ізолейцин |
40 |
Аденозін-трифосфат |
|
10 |
|
|
|
натрія |
|
|
|
DL – Валін |
50 |
Аденілова кислота |
|
0,2 |
|
DL – Глутамінова |
140 |
D – Рибоза |
|
0,5 |
|
кислотамоногідрат |
|
|
|
|
|
DL – Аспаргінова |
60 |
D – Дезоксирибоза |
|
0,5 |
|
кислота |
|
|
|
|
|
DL – α – Аланін |
50 |
Інші речовини |
|
мг |
|
L – Пролін |
40 |
Fe (NO3)3x9H2O |
|
0,1 |
|
L – оксипролін |
10 |
СН3СOONa x 3H2O |
|
50 |
|
L – Гліцин |
50 |
L – Глутатіон |
|
0,05 |
|
|
|
111 |
|
|
|
L – Цистин |
|
20 |
Глюкоза |
1000 |
||
L – Тирозин |
|
40 |
L – Глутамін |
100 |
||
L |
– |
Цистеїн |
– |
0,1 |
Феноловий червоний |
20 |
гідрохлорид |
|
|
|
|
||
Вітаміни |
|
мг |
Спирт етиловий |
16 |
||
Нікотинова кислота |
0,025 |
|
|
|||
Амід |
нікотинової |
0,025 |
|
|
||
кислоти |
|
|
|
|
|
|
Піридоксин |
– |
0,025 |
|
|
||
гідрохлорид |
|
|
|
|
||
Піридоксаль |
– |
0,025 |
|
|
||
гідрохлорид |
|
|
|
|
||
Тіамін |
|
– |
0,01 |
|
|
|
гідрохлорид |
|
|
|
|
||
Рибофлавін |
|
0,01 |
|
|
||
Пантотенат кальція |
0,01 |
|
|
|||
Інозит |
|
|
0,05 |
|
|
|
n-Амінобензойна |
|
0,05 |
|
|
||
кислота |
|
|
|
|
|
|
Холін-хлорид |
|
0,5 |
|
|
||
Біотин |
|
|
0,01 |
|
|
|
Α |
– |
Токоферол- |
0,01 |
|
|
|
фосфат натрія |
|
|
|
|
||
Фолієва |
кислота |
0,01 |
|
|
||
кристалічна |
|
|
|
|
||
Кальциферол |
|
0,1 |
|
|
||
Менадіон |
|
0,01 |
|
|
||
Вітамін |
|
А |
0,1 |
|
|
|
кристалічний |
|
|
|
|
||
Аскорбінова |
|
0,05 |
|
|
||
кислота (вітамін С) |
|
|
|
|||
У всі поживні середовища та деякі сольові розчини для визначення рН додають індикатор – феноловий червоний (в концентрації 0,002 %, яка не має токсичного впливу на клітини та віруси). Додавання індикатора дозволяє визначити момент закислення поживного середовища продуктами метаболізму клітин до рівня, який потребує заміни поживного середовища на нове (рН<7; середовище забарвлюється в жовтий колір). При
112
підвищенні рН розчин набуває червоно-малинового кольору. При нейтральному рН (7,2-7,4) колір середовища жовтогарячий. Для регулювання рН сольових розчинів та поживних середовищ використовують 7,5%-ий розчин бікарбонату натрію та 3%- ий розчин оцтової кислоти.
Усі поживні середовища за функціональним призначенням можна поділити на ростові та підтримуючі. Ростові поживні середовища забезпечують існування та розмноження клітин і містять 2-10 % сироватки крові. Ці поживні середовища застосовують, як правило, в перші дні культивування клітин. Підтримуючі поживні середовища забезпечують життєдіяльність клітин, але не їх розмноження. Вони не містять сироватки крові. Їх використовують для культивування клітин після зараження їх вірусами.
Контамінація культур клітин
Вірусологічні дослідження, які проводяться з використанням клітинних культур, потребують постійного контролю присутності сторонніх мікроорганізмів (контамінантів) – вірусів, бактерій, грибів, мікоплазм та клітин інших клітинних культур. На відміну від організму людини, імунна система якого володіє багатьма різноманітними механізмами захисту від інфекцій, клітини в культурі in vitro захищені лише обережністю роботи дослідника.
Найпоширенішими контамінантами клітинних культур є мікоплазми. І хоча культури первинних клітин не забруднені мікоплазмами, багато ліній і штамів постійних клітин мають таке забруднення. Джерелом мікоплазм можуть бути сироватка тварин, яка частіше за все буває забруднена Acholeplasma laidlawii та Microplasma arginini, а також власне дослідники, оскільки M. hominis, M. рharingis та M. salivarium легко виділяються з порожнини рота та глотки людини. Забруднення культур клітин мікоплазмами не є таким очевидним, як бактеріями. У зв’язку з цим їх своєчасне виявлення – важлива умова підтримання високої якості культури клітин. Крім того, невиявлені та не видалені мікоплазмені інфекції, як і інфекції, викликані іншими контамінантами, можуть змінювати метаболізм клітин, викликати хромосомні порушення, впливати на сприятливість клітин до вірусів, що досліджуються. Особливо це стосується латентних інфекцій.
113
Попередити розмноження та знищити бактерії, мікоплазми та гриби вдається за допомогою протимікробних препаратів (антибіотики та ін.), які додаються в ростові середовища безпосередньо перед їх використанням. Однак при тривалому культивуванні клітинних культур застосування антибіотиків не бажано. Тому при роботі з клітинами необхідно дотримуватися стерильних умов роботи для запобігання забруднення мікроорганізмами, та проводити адекватні тести для оцінки їх ефективності.
Взаємодія вірусів із клітинами
При взаємодії вірусу з клітиною можна виділити такі основні стадії:
1. Початковий період
1.1.Адсорбція віріону на поверхні клітини (зворотна та незворотна адсорбція) – прикріплення вірусних часток до клітинної поверхні (рис.10. (кольорова вклейка)) (не характерно для вірусів рослин).
1.2.Проникнення вірусу в клітину – за рахунок рецепторного ендоцитозу чи злиття вірусної та клітинної мембран.
1.3.Звільнення вірусного геному (депротеїнізація вірусу,
„роздягання” вірусної нуклеїнової кислоти). 2. Середній період ( „темнова”, екліпс - фаза)
2.1.Синтез ранніх (неструктурних) вірусних білків.
2.2.Синтез компонентів майбутніх вірусних часток –
реплікація вірусних нуклеїнових кислот та синтез пізніх (структурних) вірусних білків.
3. Заключний період
3.1.Формування віріонів – збірка вірусних компонентів.
3.2.Вихід віріонів із клітини.
Реакція клітин на вірусну інфекцію досить різноманітна. Запропоновано багато способів класифікації реакції клітин на зараження вірусами. Найбільш прийнятою є класифікація вірусних інфекцій на клітинному рівні, що запропонована В.М. Ждановим та А.Г. Букринською (1986). Вірусні інфекції підрозділяються на автономні та інтеграційні.
114

Автономний – тип вірусної інфекції, при якому вірусний геном реплікується незалежно від клітинного.
Інтеграційний - тип вірусної інфекції, при якому вірусний геном частково або повністю інтегрується з клітинним геномом та реплікується разом з ним.
Продуктивна вірусна інфекція завершується утворенням інфекційного потомства, на відміну від абортивної, для якої не характерне утворення інфекційних часток, або вони утворюються в значно меншій кількості, ніж при продуктивній вірусній інфекції.
Гостра вірусна інфекція – це така форма інфекції, за якої після утворення вірусного потомства клітина або гине, або видужує і не містить вірусних компонентів. При хронічній вірусній інфекції клітини продовжують продукувати вірусні частки або їх компоненти протягом тривалого часу і передають цю здатність спадково. Гостра інфекція, яка завершується загибеллю клітини (лізисом), носить назву цитолітичної, а гостра вірусна інфекція, яка безпосередньо не призводить до лізису клітини, і при якій клітина ще може функціонувати впродовж деякого часу, продукуючи вірусні частки, називається нецитолітичною.
При взаємодії вірусів із клітинами в культурі спостерігаються різні патологічні зміни, які зумовлені рядом факторів:
-особливістю реплікації віріонів;
-чутливістю клітин до вірусів;
-умовами культивування клітин (склад поживного середовища, рН середовища, температурою
культивування).
Патологічні зміни уражених вірусами клітин зумовлені специфічними та неспецифічними процесами. До неспецифічних процесів відносять процеси, обумовлені змінами деяких ланок обміну речовин, зміною проникності цитоплазматичної мембрани,
асаме:
-дезінтегративне набрякання ядер клітин – феномен збільшення розмірів ядра клітини в результаті стимуляції обміну речовин при вірусній інфекції;
-зміна метаболізму та мітотичного поділу клітини;
-цитотоксичну дію вірусів – вплив вірусів на клітину ще до
стимуляції обміну речовин.
Названі зміни відносять до неспецифічної взаємодії вірусів із клітинами, тому що вони не залежать від дози вірусу, розмірів
115
віріонів, типу клітин, характеру утворення включень, тривалості вірусної інфекції.
До специфічних процесів взаємодії вірусу з клітиною відносять:
-різноманітні зміни, що проявляються в пригніченні синтетичних процесів, порушенні функціональної активності, пошкодженні структур клітини та її загибелі (ЦПД, утворення включень);
-цитопроліферативний ефект, який, як правило, призводить до трансформації клітин;
-резистентність клітин до вірусів;
-вірусопатичну дію;
-синтез неповноцінних віріонів та вірусних специфічних антигенів (при абортивній інфекції);
-індуктивний вплив вірусу (синтез інтерферонів).
Методи виявлення та ідентифікації вірусів у клітинних культурах
Існують наступні основні методи індикації (виявлення) вірусів у культурі клітин:
-за цитопатичним ефектом чи цитопатичною дією (ЦПЕ,ЦПД);
-за виявленням внутрішньоклітинних включень;
-електронною мікроскопією;
-в реакціях імуноферментного та радіоімунного аналізу (ІФА, РІА);
-в реакції гемадсорбції (РГАд) (рис.15 (кольорова вклейка));
-в реакції імунофлюоресценції;
-за виявленням інтерференції вірусів;
-за пригніченням метаболізму клітин (кольорова проба);
-за утворенням бляшок.
Цитопатична дія
Найбільш широко та часто про розмноження вірусу в культурі клітин роблять висновок за цитопатичним ефектом чи цитопатичною дією (ЦПЕ, ЦПД). ЦПД – це явище, що призводить до руйнування структури клітин під впливом вірусів.
116
Причини ЦПД:
-порушення нормальної життєдіяльності клітин в результаті механічного пошкодження клітинних структур вірусними компонентами (дефекти цитоплазматичної мембрани, які виникають в результаті проникнення чи виходу вірусів із клітини);
-руйнування лізосом і вихід їх ферментів у цитоплазму (автоліз клітин);
-виснаження білкових та енергетичних ресурсів клітин за рахунок “переключення” клітинних ферментів та білоксинтезуючого апарату на синтез вірусспецифічних компонентів;
-порушення клітинного моношару.
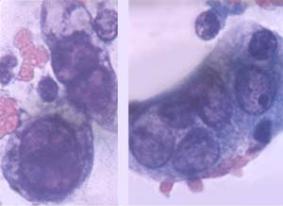
За терміном виникнення ЦПД поділяють на ранню та пізню. Рання ЦПД виявляється в перші години після інфікування (від 3 годин після інфікування клітинних культур вірусом), спричиняється дією структурних елементів вірусів/ У цей період ще не відбулося проникнення вірусу в клітину. Даний тип ЦПД морфологічно проявляється в порушенні клітинного моношару, заокругленні клітин, відокремленні їх від скла(рис.2.23.а.).
Пізня ЦПД виявляється, коли вірус (чи його геном) потрапляє в клітину і, як правило, на кінцевих стадіях репродукції вірусів у клітині. Пізня ЦПД проявляється в утворенні полікаріоцитів і включень (рис.2.23.b.).
a.На ранній стадії розвитку інфекції уражені клітини епітелію збільшені у розмірах, звичайно містять одне ядро, що має вигляд часового скла,
оскільки хроматин розташовується по переферії ядра, а центральна
. частина останнього залишається світлою, вільною від від хроматину. Ядерця збільшені у розмірах.
117
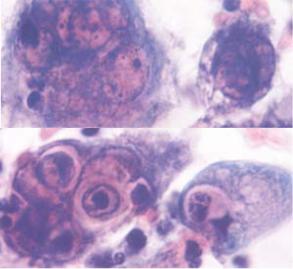
b.Протягом пізньої стадії захворювання, а також при рецидиві інфекції, виявляються багатоядерні клітини ураженого епітелію. Ядра містять включення – комплекси вірусних часток. Навколо ядер спостерігається освітлення цитоплазми.
Рис.2.23. Рання та пізня ЦПД при герпесвірусній інфекції
Для характеристики деструктивних змін одношарових культур клітин, що заражені різними вірусами, найчастіше використовується робоча класифікація ЦПД (О.Г. Анджапарідзе, 1962):
1 група. Рівномірно розподілена дрібнозерниста деструкція клітин – виникає при ураженні клітин вірусами поліомієліту, Коксакі, ЕСНО (рис.2.24.a, рис.11.a. (кольорова вклейка)), ЕСМО, віспи, грипу та ін.
2 група. Вогнищева дрібнозерниста деструкція клітин з тяжами незмінених клітин між ними - виникає при ураженні клітин вірусами весняно-літнього енцефаліту, вірус хвороби Ауескі (псевдосказу), спумавірусами (рис.2.24.b) та ін.
3 група. Осередкові скупчення заокруглених клітин, що нагадують грона винограду - виникає при ураженні клітин аденавірусами (рис.12 (кольорова вклейка)).
4 група. Рівномірно-розподілена крупнозерниста деструкція клітин (клітини збільшені у розмірі, заокруглені) - виникає при ураженні клітин вірусом простого герпесу (рис.11.b. (кольорова вклейка)), мавп’ячим вірусом В та ін.
5 група. Утворення гігантських багатоядерних клітин – симпластів та синцитіїв - виникає при ураженні клітин вірусами кору, вісповакцини, герпесу та ін (рис.13 (кольорова вклейка)). При цьому типі ЦПД відбувається розчинення клітинних оболонок, внаслідок чого цитоплазма сусідніх клітин зливається,
118

утворюючи єдине ціле, в якому (в основному по периферії) розташовані ядра клітин.
a |
b |
Рис.2.24. Заокруглення клітин у випадку зараження культури клітин вірусом ECHO (a) та цитопатична дія спумовірусу на клітини (b)
Слід розрізняти термін симпласт та синцитій. Різниця між цими поняттями полягає в тому, що синцитії виникають у результаті часткового злиття цитоплазматичної мембрани клітин, на відміну від повного злиття мембрани при утворенні симпластів. Таким чином, синцитій – це група клітин, які з’єднані між собою протоплазматичними відростками, а симпласти мають загальну масу протоплазми, в якій міститься багато ядер.
Внутрішньоклітинні включення
При багатьох вірусних інфекціях в клітинах (в ядрі чи цитоплазмі) різних органів та тканин з’являються особливі утворення, які називають тільцями-включеннями. Їх класифікують за локалізацією в клітині, вмістом нуклеїнової кислоти, тинкторіальними властивостями та гомогенністю (гомогенні чи зернисті).
Тільця-включення локалізуються вибірково. Так, цитоплазматичні включення характерні при віспі (тільця
119
Гварнієрі), сказі (тільця Бабеша-Негрі) (рис.14.a. (кольорова вклейка)), грипі, парагрипі, чумі великої рогатої худоби та ін. При ринотрахеїті великої рогатої худоби, ларинготрахеїті птахів, аденовірусній інфекції(рис.14.b. (кольорова вклейка)), герпесвірусній інфекції (тільця Каудрі), ящурі розвиваються включення в ядрі.
По відношенню до барвників включення поділяють на базофільні (фарбуються основними барвниками – азур, піроксин) та ацидофільні або оксифільні включення (фарбуються кислими барвниками – еозин, кислий фуксин).
За своєю природою включення можуть бути місцями утворення вірусних часток (фабрики віріонів – тільця Гварнієрі), скопиченням вірусів (тільця Бабеша – Негрі, поліедри), глибками хроматину (тільця Каудрі).
Титрування вірусів у одношарових культурах клітин
У лабораторних роботах з вірусами, біофабричному виробництві та у медичній і ветеринарній практиці постійно виникає необхідність визначення кількості вірусів у тому чи іншому матеріалі. Без такого визначення неможливе контрольоване експериментальне ураження вірусами лабораторних тварин, виробництво живих та інактивованих противірусних вакцин та діагностичних препаратів, оцінка активності живих противірусних вакцин, отримання імунних сироваток та багато інших завдань.
Кількість вірусу в будь-якому матеріалі визначають за титром вірусу в цьому матеріалі. Під титром вірусу розуміють вираження його концентрації у матеріалі. Титр вірусу – це кількість вірусу, яка міститься в одиниці об’єму матеріалу.
Титрування вірусів та антитіл до них в одношарових культурах клітин проводиться наступними методами: за ЦПД, методом кольорової реакції (відбувається зміна забарвлення у жовтий колір середовища під впливом продуктів метаболізму неуражених клітин, в той час як у тканинній рідині заражених вірусами клітин зберігається вихідний колір середовища - червоний), методом бляшок (рис.15 (кольорова вклейка)), у реакції гемадсорбції (рис.16 (кольорова вклейка)). Оскільки кількість вірусу неможливо виразити у звичайних одиницях об’єму, маси та ін., застосовують вимір одиниць дії чи одиниць активності
120