

Lektsii_po_fiziki
.pdfЗа счет указанных белков полностью или частично осуществляются специфические функции мембран: проницаемость, активный транспорт через мембрану, генерация электрической разности потенциалов и др.
Мембраны не являются неподвижными структурами. Липиды и белки обмениваются местами и перемещаются вдоль плоскости (латеральная диффузия) и поперек плоскости мембран (диффузия флип-флоп) .Латеральная диффузия сопровождается высокой подвижностью липидов, а диффузия флип-флоп – низкой подвижностью, т.е. обмен местами липидов, находящихся на разных сторонах мембраны, является маловероятным процессом.
Физиологические функции мембран.
1)Делят клетку на отдельные участки, фазы, в которых протекают различные биохимические реакции, которых в клетках протекает до тысячи видов.
2)Принимают участие во всех процессах обмена веществ, которые обусловливают жизнедеятельность клетки. Это происходит потому, что мембранные структуры в организмах составляют огромную поверхность (до десятков тысяч квадратных метров). Поэтому биохимические реакции протекают в основном на поверхности мембран, обеспечивая обмен веществ.
3)Координируют и регулируют биохимические и биофизические процессы в клетках. Мембраны являются своеобразным устройством, воспринимающим сигналы извне и преобразующим их в команды, регулирующие обмен веществ в клетке.
4)Клеточные мембраны обеспечивают адгезию (сцепление) клеток друг с другом, что обеспечивает существование ткани. Адгезия осуществляется специализированными структурами на клеточных мембранах (иногда это просто как механическое сцепление типа”гнездо-шип", иногда химическая связь, иногдасклеивающее цементнообразное вещество”).
2.Физические свойства мембран.
1. Измерение подвижности молекул мембраны и диффузия частиц через мембрану свидетельствуют о том, билипидный слой ведет себя подобно жидкости. Но, с другой стороны, мембрана есть упорядоченная структура, свойственная кристаллам. Эти два факта заставляют думать, что фосфолипиды в мембране находятся в жидкокристаллическом состоянии.
81

2.Вязкость липидного слоя мембраны на 2 порядка (в 100 раз) выше, чем у воды. От вязкости липидного слоя зависит скорость поступательного и вращательного движения молекул (в том числе мембранных белков и ионов), т.е. вязкость определяет подвижность ионов и молекул, проникающих через мембрану (проницаемость мембраны), а, следовательно, регулирует скорость протекающих в мембранах ферментативных реакций.
3.Поверхностное натяжение липидного слоя на 2-3 порядка (100-1000 раз) меньше, чем у воды. Значит поверхностная энео-
гия липидного слоя незначительна фузию веществ через мембрану.
æ |
σ = |
WП ö |
, что облегчает диф- |
|
è |
S |
ø |
||
ç |
÷ |
|
||
4.При изменении температуры в мембране наблюдаются фазовые переходы: плавление липидов при нагревании и кристаллизация при охлаждении. Структура молекул в жидком и кристаллическом состояниях различна. В жидкой фазе молекулы фосфолипидов могут образовывать полости (кинки), в которые способны внедряться молекулы диффундирующего вещества.
5.Двойной фосфолипидный слой уподобляепт мембрану конденсатору. Электроёмкость 1 мм2 мембраны составляет 5-13 нФ.
Модели искусственных мембран для изучения свойств мембран.
1.Частокол Ленгмюра – монослой фосфолипидов. Модель создана в 1917 году.
На поверхность воды наносят каплю растворенных в каком-либо растворителе фосфолипидов или жирных кислот. После распределения их молекул на поверхности воды и испарения расворителя на поверхности воды остается пленка. Ад-
сорбированные молекулы липидов располагаются перпендикулярно поверхности воды.
2. Липосомы – широко распространенная модель. Это мельчайшие пузырьки, состоящие из билипидной мембраны. Получаются при обработке смеси воды и фосфолипидов ультразвуком. На ней хорошо изучать влияние состава фосфолипидов на свой-
82

ства мембран или влияние мембран на свойства втсраеваемых белков.
3. Билипидная мембрана. Берется два водных раствора, разде-
ленных тефлоновой перегородкой с отверствием. Отверствие заполняют фосфолипидом, растворенным в гексане. Когда растворитель и изли-
шек липида растекаются по тефлону, в отверстии образуется бислой, тол-
щиной в несколько нанометров.
На такой мембране удобно изучать проницаемость мембраны для разных агентов, измерять сопротивление мембраны или генерируемый на ней потенциал.
Методы исследования мембран
1. Микрокалориметрия. Изучаются фазовые переходы на каком-либо участке мембраны и оценивают размеры “кооперативных единиц”, т.е. число молекул, участвующем в фазовом переходе. Для этого измеряют теплоемкость C суспензии фосфолипидов при разных температурах в области фазового перехода. Например, для синтетического липида дистеароил-фосфати- дил-холина графики зависимости C от T , где T - абсолютная температура выглядят так
83

C = QT
В области фазового перехода при TC происходит резкое возрастание теплоемкости. Площадь под кривыми соответствует количеству теплоты Q , поглощаемого при переходе из твердого состояния в жидкое. Зная массу m фосфолипида в пробе, можно рассчитать энтальпию плавления
H = mQ
и энтропию при плавлении
S = TH .
C
Энтрапия S возрастает с ростом длины углеводородной цепи жирных кислот, причем на каждую метиленовую группу приходится примерно постоянное увеличение энтропии при фазовом переходе. Т.о. мы можем судить о длине куглеводородной цепи липидов.
Площадь под кривой от T1
S = Tò2CdT
T1
пропорциональна количеству молекул уже претерпевших фазовый переход.
2. ИК - спектроскопия.
84
Этот метод основан на замечательном свойстве природы: молекулы каждого вещества имеют индивидуальные, специфические спектры поглощения. Молекулярные спектры позволяют исследовать состав мембран, строение молекул в мембране и характер межмолекулярных взаимодействий.
3.Люминесцентный анализ.
Производят на спектрофлюоиметрах.
Молекулы белков обладают флюоресценцией. Параметры флюоресценции чувствительны к структуре окружения флюоресцирующей молекулы. Поэтому по люминесценции можно изучать химические превращения и межмолекулярные взаимодействия.
Для исследования мембран к мембранным системам добавляют флюоресцирующие молекулы: флюоресцентные зонды, если молекула образует нековалентную связь с мембраной , или флюоресцентные метки, если молекула образует химическую связь с мембраной.
При освещении мембранной системы с флюоресцирующей молекулой поляризованным светом люминесценция оказывается также поляризованной. Если флюоресцентный зонд неподвижен (например, раствор заморожен), то степень поляризации P0 будет мексимальной. Однако, если зонд или метка находятся в жидком растворе, то он за время возбужденного состояния успевает переориентироваться (перестроится), а поэтому изменится и степень поляризации люминесценции до величины P . Изменение степени поляризации зависит от вязкости окружающих зонд молекул. Т.о люминесцентный анализ позволяет определить вязкость мембран, а также конформационные перестройки в белках
имембранах.
4.Рентгеноструктурный анализ.
В основе этого метода лежит дифракция рентгеновского излучения на структуре мембран. Т.к. липиды мембраны есть жидкий кристалл, то дифракционная картина непосредственно связана с внутренней структурой кристалла, т.е. с расположением атомов в кристалле. Т.о. рентгеноструктурный анализ позволяет определить строение и параметры липидных молекул.
5.ЭПР
6.ЯМР
85

К методу рентгеноструктурного анализа мы вернемся при изучении рентгеновского излучения, а методы ЭПР и ЯМР мы изучим в конце года.
Диффузия в жидкостях. Закон Фика
Важным элементом функционирования мембран является их способность пропускать или не пропускать атомы (молекулы) и ионы, т.е. проницаемость мембран.
Вероятность такого проникновения частиц зависит как от направления их перемещения (например, в клетку или из клетки), так и от разновидности молекул и ионов.
Эти явления изучаются в разделе физики, называемолм явлениями переноса (диффузия, теплопроводность, перенос импульса, электропроводность). Эти явления представляют собой необратимые процессы.
Мы рассмотрим только те явления, которые характерны для мембран: диффузию и перенос заряда (синоним слова перенос в биофизике - транспорт).
Диффузия – самопроизвольный процесс проникновения вещества из области большей концентрации в область меньшей концентрации в результате теплового хаотичного движения.
Качественными характеристиками диффузии являются:
1.Поток вещества
Φ= mt - масса вещества, переносимого через площадь S , перпен-
дикулярную движению частиц, в единицу времени.
2. Плотность потока вещества I = ΦS = S mt - масса вещества, пере-
носимого через единицу площади S , перпендикулярной движению частиц, за единицу времени.
Молекулы жидкости перескакивают из одного равновесного состояния в другое. В одном из равновесных состояний поместим площадку S и определим, какое число молекул пройдет через неё из двух ближайших равновесных состояний 1 и 2.
86

Отложим слева и справа от S расстояния δ (длина свободного пробега молекулы) и построим параллепипеды 1и 2, площадью S и толщиной l << δ .
Объёмы параллепипедов V1 = V2 = S × l .
Для упрощения будем считать, что молекулы движутся с одинаковыми средними скоростямиυ = δτ , где τ - время свободно-
го пробега (перескока).
Пусть число молекул в единице объёма (концентрация) в первом объёме n1, во втором n2 . Тогда общее число молекул в первом объёме n1 × S × l , во втором - n2 × S × l .
Молекулы движутся хаотично по всем трем направлениям: x, y, z . Из них 13 движется в направлении оси Ox, 13 - Oy , 13 - Oz
. Причем в направлении x , например, половина из этой 13 движется влево, половина - вправо. Поэтому вдоль оси Ox от перво-
го параллепипеда к площадке S |
движется |
1 |
n1Sl молекул, а от |
||
6 |
|||||
второго - |
1 |
n2 Sl . Т.о. за время |
t через площадку S проходит |
||
6 |
|||||
число молекул |
|
|
|
||
N = 16 n1Sl − 16 n2 Sl = 16 Sl(n1 − n2 ).
Пусть масса одной молекулы m0 , тогда за время t через площадку S переносится масса вещества
87

Dm = m0 × DN = 16 m0Sl(n1 - n2 ) .
Время пролета молекулами площадки S параллепипедов 1 и 2 (через центральную оно будет таким же)
t = υl = lδ×τ .
Поток вещества через площадку определится как
F = Dmt = 16 × mD0Slt (n1 - n2 ) = 16 × m0τSδ (n1 - n2 ) .
Можно показать, что
n1 - n2 = dndx × 2δ ,
где dndx - градиент концентраций. Поток вещества тогда запишем как
F = |
1 |
× |
m Sδ |
× |
dn |
× 2δ = |
1 |
× |
m Sδ 2 |
× |
dn |
6 |
0 |
dx |
3 |
0 |
dx |
||||||
|
|
τ |
|
|
|
τ |
|
Через единицу площади переносится поток вещества (плотность потока)
I = F = 1 × m0δ 2 × dn .
S 3 τ dx
Введем массовую концентрацию c = m0n . Тогда
m0 dndx = dmdx0n = dcdx - градиент массовой концентрации.
Обозначим |
1 |
δ 2 |
и назовем её коэффициентом диффузии, |
3 |
τ = D |
а также учтем, что суммарная плотность потока направлена в сторону уменьшения концентрации, и укажем это напрвавлние знаком “-“. Получим уравнение Фика:
I = -D dcdx .
[D] =1 мс3 . Физический смысл коэффициента диффузии: он числен-
но равен массе вещества, переносимого через единичную площадку за 1 секунду при градиенте концентраций равном 1.
[I ] = 1 |
кг |
|
, если [с] =1 |
кг |
, |
||||
|
2 |
3 |
|||||||
|
|
с × м |
|
|
|
м |
|
|
|
[I ] = 1 |
|
моль |
, если [с] =1 |
|
моль |
||||
с × м |
2 |
|
м |
3 |
|||||
|
|
|
|
|
|
|
|||
88
Лекция 7.
Транспорт веществ через мембрану.
Являясь открытой термодинамической сиситемой, клетка постоянно осуществляет обмен веществом с окружающей средой. Такой обмен возможен благодаря способности клеток пропускать различные вещества через свою оболочку. Эта способность клеток называется проницаемостью.
Перемещение веществ в клетку или из неё в окружающую среду может осуществляться многими способами. В зависимости от того, что является источником энергии для переноса вещестыва, что является силой перемещения, все виды переноса веществ можно разделить на пассивный и активный транспорт.
Пассивный транспорт веществ.
Пассивный транспорт всегда осуществляется за счет энергии, сконцентрированной в каком-либо градиенте. Энергия метаболических процессов клеток (энергия гидролиза АТФ) на этот процесс непосредственно не переносится. Пассивный транспорт всегда идет от более высокого энергетического уровня к более низкому.
Основными градиентами, присущими живым организмам, являются градиенты концентрационные, электрические, осмотические, градиенты гидростатического давления.
В соответствии с этими градиентами имеются следующие виды пассивного транспорта веществ в клетках и тканях: диффузия, осмос, электроосмос, аномальный осмос, фильтрация.
Основным механизмом пассивного транспорта является диффузиясамопроизвольный процесс проникновения вещества из области большей концентрации в область ментшей концентрации в результате теплового хаотического движения.
Перенос незаряженных частиц (атомов и молекул) через мембрану
Очевидно, что при переносе незаряженных частиц единственным градиентом является градиент концентраций. Сле-
89
довательно, механизм переноса молекул и атомов через мембрану
– диффузия, а сам транспорт описывется уравнением Фика:
I = −D dcdx
Концентрационный градиент клеточной мембраны определить трудно, поэтому лучше использовать более простое уравнение, предложенное Коллендором и Берлундом и которое легко вывести из уравнения Фика.
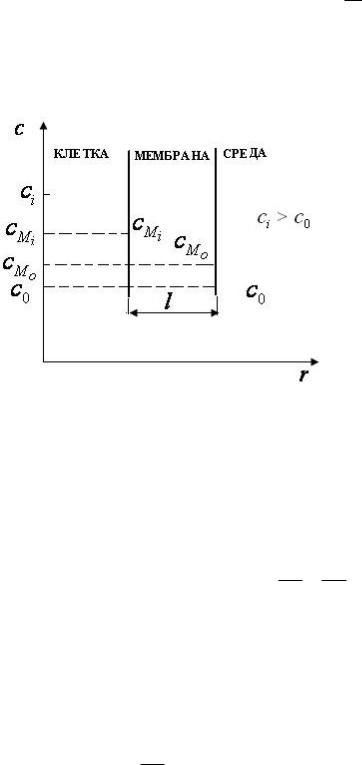
ci - концентрация вещества в
клетке;
c0 - концентрация этого же
вещества снаружи клетки; cMi - концентрация этого же
вещества внутри клетки на границе клетка - мембрана; cMo - концентрация этого же
вещества внутри клетки на границе окружающая средамембрана.
ci > c0 . Следовательно, перенос идет из клетки в окружающую среду. Для мембраны
dc |
= |
c2 |
− c1 |
= |
cMo − cMi |
. |
dx |
|
l |
l |
|||
|
|
|
|
Измерить cMi и cMo трудно, но зато экспериментально можно измерить ci и c0 . Кроме того, считают, что
cMi = cMo = k
ci co
k - коэффициент распределения частиц между мембраной и окружающей средой. Следовательно,
|
cMo = kco ; |
cMi = ci |
|
|
|
|
Тогда |
|
|
|
|
||
I = −D |
cMo − cMi |
= − |
Dk(co − ci ) |
= |
Dk(ci − co ) |
. |
|
l |
|
l |
|
l |
|
Обозначим P = Dkl - проницаемость мембраны, получим
I= P(ci − co )
-это и есть уравнение Коллендора-Берлунда.
90