

5. Особенности русского генофонда
Российские ученые исследовали шесть массивов данных - два антропологических (соматология и дерматоглифика), два массива ДНК полиморфизма (мтДНК и Y- хромосомы), классические генетические маркёры и географию русских фамилий. Целью исследования было выяснить структуру русского генофонда. Средством стали методы геногеографии.
Соматология. Антропологические данные описывают физический облик русского населения. Впервые проанализирован весь комплекс данных по антропологии коренного русского населения в пределах «исконного» ареала (Русской равнины). Суммарно изучено 180 популяций по 18 признакам соматологии: длина тела, цвет глаз и волос, форма спинки носа и уплощенности лица, рост бороды и другие.
Некоренное население и русские популяции за пределами «исконного», исторического ареала (например, Урала, Сибири, Поволжья) не учитывалось.
«Простые» карты. Рассматривая карты отдельных признаков, видишь, что география большинства из них довольно запутана. Однозначные тренды обнаружены лишь для немногих признаков. Изменчивость всех 18 признаков мы обобщили в картах главных сценариев (канонических переменных) и картах генетических расстояний.
Главный сценарий. Карта первой канонической переменной выявляет широтную изменчивость. Этот статистический метод позволяет выявить то основное, что присутствует на картах всех отдельных признаков. Значит, антропологический облик русских популяций меняется главным образом при движении с севера на юг (или, то же самое, с юга на север). Такую же закономерность хорошо знают и лингвисты - они выделяют северные, южные и смешанные среднерусские говоры. Итак, различия между русским севером и русским югом - это первостепенное различие внутри русского населения. Но ни о какой границе между севером и югом не может быть и речи - изменения происходят очень постепенно. Они скорее задают главную ось, вдоль которой мы видим наибольшую изменчивость.
Генетические расстояния. Иную, но не менее важную информацию несёт обобщённая карта второго типа - карта генетических расстояний. На ней показано, насколько каждая из русских популяций отличается от среднестатистической русской популяции. Популяции, похожие на эту «среднюю русскую», встречаются повсюду, но в основном сосредоточены в центре русского ареала. Из предыдущей карты можно было бы ожидать что «самые отличающиеся» расположатся одни на юге, другие на севере. Но реальная картина иная. Популяции, значительно отличающиеся от средних русских характеристик, образуют на карте скопления, ядра. Одна группа этих ядер расположена на западе, другая - в восточной части «исконного» русского ареала.
Для объяснения этой картины вспомним, что современное русское население образовалось при смешении пришлого славянского и автохтонного финно-угорского населения. Славянские племена в течение нескольких веков продвигались с запада на восток, колонизируя Восточно-Европейскую равнину и ассимилируя местные финно-угорские племена. В середине этого процесса, когда славяне достигли территорий, которые являются сейчас серединой русского ареала, смешение было особенно интенсивным. В результате образовался «среднерусский» генофонд, составленный из славянского и финно-угорского компонентов. И на нашей карте как раз в середине ареала мы видим «среднестатистический» русский тип. В конце процесса славянской колонизации, в восточных областях, местное финно-угорское население численно преобладало над пришлым славянским. И на нашей карте на востоке мы видим отличия от «среднестатистического» типа - за счёт преобладания финно-угорского компонента. А в начале того же процесса, в западных областях, численно преобладало славянское население. И на нашей карте на западе мы видим отличия от «среднестатистического» типа - но теперь за счёт преобладания славянского компонента. Предлагается гипотеза: в современном русском генофонде разных территорий видны три фазы славянской колонизации: преобладание на западе славянского, в центре смешанного и на востоке дославянского (в основном финно-угорского) населения.
Эта гипотеза подтверждается тем, что зона «среднестатистического» русского населения соответствует продвижению границ Русского государства (при расширении его пространства) на восток в IX-XI веках - то есть соответствует той зоне, куда передвигались наибольшие массы славянского населения и где можно ожидать наибольшего смешения с финно-угорскими популяциями. Второе подтверждение: западные «ядра» на антропологической карте совпадают с ареалами летописных славянских племён на археологической карте: каждое из западных ядер можно сопоставить с конкретным славянским племенем.
Однако это направление «запад<=>восток» так и не стало главным сценарием русского генофонда, не сумело превозмочь различия между севером и югом. На севере же остаются крупные «залежи» дославянского генофонда. Генофонд Русского Севера нельзя рассматривать только как наследство от финно-угров: он часто тяготеет к западным территориям, и скорее всего, к самому древнему палеоевропейскому пласту генофонда Европы. Когда с помощью генетики будет прослежена цепь поколений вплоть до первых основателей европейского генофонда, то, скорее всего, окажется, что этот пласт удалось сохранить не только современным финно-уграм, но и балтам, и славянам.
Антропология: кожные узоры. Был нанесен на карту и другой класс антропологических признаков - дерматоглифических. Они охватывают меньшее число популяций «исконного» русского ареала - 28. И самих признаков меньше - всего 7, но зато это избранники. Эти семь признаков носят звание ведущих: они не коррелируют друг с другом и несут основную информацию о кожных узорах.
«Простые» карты. Карты отдельных признаков опять оказались разнообразны. Особенно любопытно распространение европеоидно-монголоидного комплекса - это такое сочетание признаков кожных узоров, которое хорошо дифференцирует европеоидные и монголоидные популяции. Ожидалось, что в русском населении монголоидность будет возрастать с запада на восток. Но нет, на карте вообще не видно никаких закономерностей - то есть в пределах «исконного» русского ареала распределение дерматоглифического европеоидно-монголоидного комплекса хаотично. Т.е., по этому признаку, татарско-монгольское иго на русском генофонде никак не сказалось.
Главный сценарий. Зато обобщённая карта (по всем дерматоглифическим признакам) выявляет чёткую закономерность. Обобщение провели методом главных компонент, широко распространённым в геногеографии. Этот метод аналогичен методу канонических переменных, который был применен к антропологическим данным и обнаружил у них широтную изменчивость. И теперь, уже по данным дерматоглифики, первая главная компонента также выявила постепенную широтную изменчивость в русском населении.
Итак, изучение двух независимых комплексов признаков единодушно засвидетельствовало, что главный компас антропологии указывает с юга на север: в этом направлении изменяется русский фенофонд, то есть совокупности внешних признаков - фенов. А что же гены, что же собственно генофонд, что скажет генетика?
Генетика: классические генные маркёры. Классические маркёры генетики - это все те гены, которые популяционная генетика использовала до «ДНК эры» - например, группы крови и гены ферментов. Классические маркёры изучались на протяжении десятилетий. Созданный в РАН банк данных «Русский генофонд» включает почти всё, что было опубликовано по русскому населению. Были построены 66 карт аллелей 24 локусов, но в анализ главных компонент были включены только наиболее изученные (35 аллелей 13 локусов). Каждый из генов изучен в разном числе популяций, в среднем - по 35 популяциям. Напомним, что антропологические признаки были изучены в пять раз подробнее - по 180 популяциям.
«Простые» карты. Карты отдельных генов выявили сложные картины - как это было и для отдельных признаков антропологии и дерматоглифики. Хотя опять, для части генов видны чёткие закономерности. Например, частота аллеля НР*1 постепенно убывает с юго-запада к северу, АСР1*А убывает с юго-востока к северо-западу, а частота GL01*1 максимальна в центре русского ареала и снижается к периферии. Частота аллеля А системы групп крови АВО высока на западе и снижается к востоку. А вот ген группы крови резус (RH*d), как и большинство прочих генов, не обнаруживает простой закономерности в русском ареале: области высоких и низких частот чередуются на карте либо беспорядочно, либо образуют сложную, неоднозначную мозаику.
Главный сценарий. Для обобщения всего этого разнообразия отдельных карт были построены карты главных компонент. И первая главная компонента выявила широтную изменчивость. Вновь - теперь в изменчивости классических маркёров - эта закономерность оказалась главной для русского генофонда.
Гетерогенность. По классическим маркёрам проведён и другой вид анализа - межпопуляционной изменчивости. Эта мера гетерогенности русского генофонда показывает, насколько разные русские популяции отличаются друг от друга. Такие показатели гетерогенности рассчитали и для многих других народов. Оказалось, что все западноевропейские народы в целом гомогенны (разные популяции, например, французов генетически очень похожи друг на друга), а сибирские народы, напротив, гетерогенны (разные популяции, например, якутов очень различаются между собой). Народы Восточной Европы, Кавказа, Урала, заняли промежуточное положение (умеренная гетерогенность). Учитывая большие различия между региональными «стандартами» гетерогенности, сравнили русский народ с народами того же самого региона - Восточной Европы. На восточноевропейском фоне, где средняя гетерогенность этноса составила лишь GST=0.7) гетерогенность русского народа велика (GST=2.0). На большом «исконном» ареале русского генофонда было неудивительно обнаружить большое генетическое разнообразие русских популяций.
Генетика: митохондриальная ДНК. Этот особый тип генетических маркёров (однородительских ДНК маркёров, то есть передаваемых из поколения в поколение только по одной линии - материнской) сейчас наиболее популярен во всём мире. Но изучение русских популяций началось недавно, имеются надёжные данные только по семи русским популяциям.
Генетические расстояния. Такое небольшое число популяций не позволяет провести надёжный анализ картографическими методами. Статистическими методами выявляется один компактный кластер, включающий большинство русских популяций, и в отдалении находится другой кластер самых северных популяций.
Гетерогенность. Величина межпопуляционных различий составила 3.3. Гетерогенность других народов Западной и Восточной Европы обычно заметно меньше (например, гетерогенность украинцев по мтДНК в пять раз меньше), то есть по митохондриальной ДНК, как и по классическим маркёрам, разные русские популяции генетически весьма различны.
Спектр. Митохондриальная ДНК позволяет провести ещё один вид анализа - по спектру гаплотипов. Если гаплогруппы мтДНК можно уподобить отдельным антропологическим или классическим маркёрам, то гаплотипы мтДНК являются очень дробными и многочисленными вариантами в пределах каждой гаплогруппы. Этим мтДНК похожа на фамилии, где мы также имеем множество вариантов одного «локуса». Можно надеяться, что многочисленность анализируемых признаков увеличивает надёжность анализа. Определили, какие гаплотипы мтДНК встречены в русских популяциях, и сравнили со спектром гаплотипов в смежных группах населения. Оказалось, что русские более всего похожи на остальные восточнославянские популяции (30% гаплотипов, обнаруженных у русских, встречены также у белорусов и украинцев). На втором месте по сходству с русскими стоят восточные финно-угорские народы, на третьем - западные финно-угорские народы, и лишь затем идут западные и южные славяне. Итак, по спектру гаплотипов мтДНК русский генофонд приближен скорее к финно-угорскому, чем к «праславянскому» генофонду, что вновь показывает роль финно-угорского субстрата в русском генофонде.
Генетика: Y- хромосома. Из всего множества генетических маркёров гаплогруппы Y - хромосомы - самый новый инструмент и, по мнению многих исследователей, самый перспективный. Они тоже однородительские ДНК маркёры: передаются из поколения в поколение только по одной линии - отцовской (как мтДНК - по материнской). Определили частоты гаплогрупп Y- хромосомы для 14 русских популяций.
«Простые» карты. Если для антропологии и классических маркёров карты отдельных признаков сложны, и чёткие закономерности выявляются лишь обобщёнными картами, то на картах отдельных гаплогрупп Y- хромосомы видны простые, чёткие картины: из восьми изученных гаплогрупп для четырёх выявлена клинальная изменчивость (постепенные изменения в одном направлении).
Главный сценарий. Первая главная компонента, обобщающая изменчивость отдельных гаплогрупп, демонстрирует широтную изменчивость.
Статистический анализ, как и в случае с мтДНК, выявляет плотный кластер центрально-южных популяций, тогда как северные русские популяции занимают каждая особое место. Степень различий между русскими популяциями (гетерогенность русского генофонда) по Y- хромосоме особенно велика - 13.6. (Например, гетерогенность по Y- хромосоме соседних украинских популяций оказалась на порядок меньше - около 1). Вспомним, что по классическим маркёрам гетерогенность русского народа была 2.0, а по мтДНК - 3.3. По этим результатам столь хорошо выявляемых различий между популяциями остаётся лишь присоединиться к мнению о перспективности Y- хромосомы для изучения русского генофонда.
Ономастика: фамилии. Если антропологические маркёры - самые изученные, иммуно - биохимические маркёры - самые классические, маркёры митохондриальной ДНК - самые популярные, Y- хромосомы - самые перспективные, то фамилии - это самые необычные маркёры. При этом они изучены подробнее, чем антропологические, имеют научный стаж лишь немногим менее чем классические и являются весьма перспективными.
Фамилии наследуются, и поэтому могут использоваться как аналоги генов. Когда мы говорим о любом признаке - мы всегда говорим о популяциях, а не об индивидах. Поэтому и фамилии рассказывают нам о закономерностях популяций, а не об отдельных людях, которые могут сменить фамилию по своему собственному усмотрению. (Ведь если Петрова станет Ивановой, а Иванова - Петровой, то в фамильном фонде популяции ничего не изменится).
Фамилии можно изучить чрезвычайно подробно. Поэтому по фамилиям можно прогнозировать генофонд там, где недостаточно времени и средств, чтобы изучать его напрямую - то есть практически везде! За несколько лет было изучено население пяти областей - фамилии миллиона человек, представляющих сельское население в пределах «исконного» русского ареала. Пять регионов - Северный, Южный, Западный, Восточный, Центральный - стали каркасом исследования. Картографирование провели только для 75 фамилий - по ним имелись данные для всего ареала. А закономерности всех десятков тысяч фамилий изучены статистическими методами по пяти «опорным» регионам.
«Простые» карты. Что же дало картографирование семидесяти пяти фамилий? Оказалось, что почти каждая фамилия имеет свой географический ареал, за пределами которого она отсутствует или крайне редка. Даже такие, казалось бы, повсеместные фамилии, как Иванов, Васильев, Смирнов, оказались отнюдь не повсеместными: Ивановых очень мало на юге, Васильевы сосредоточены в северо-западных областях, а Смирновы - в восточных и центральных.
Главный сценарий. Эти 75 карт обобщены на картах первой и второй главной компонент. Первая компонента рисует постепенные изменения от северо-запада к востоку и к югу. Эта компонента сформирована в основном «календарными» фамилиями, которые (как, например, «Васильев») сконцентрированы на западе «исконного» русского ареала. Вторая компонента показывает широтную изменчивость: различия между южными и среднерусскими популяциями, но на севере эта закономерность нарушается - северные популяции резко своеобразны. Итак, по фамилиям тоже выявили широтную изменчивость, но она осложнена, запутана, затушёвана. Возможно, эти 75 фамилий просто недостаточно представляют весь фамильный фонд.
Спектр. Поэтому статистический анализ провели по всему спектру фамилий (67 тысяч). Чтобы изучить коренное население, учли лишь деревни и небольшие города. Чтобы отсеять пришлые, «залётные» фамилии, для каждой популяции исключили те фамилии, которые встречены в районной популяции менее чем у 5 человек. Оставшиеся 14 тысяч фамилий сочли условно коренными, и работали дальше только с ними.
Оказалось, что из этих четырнадцати тысяч фамилий 250 повсеместны - встречены в каждом из пяти регионов. Остальное множество фамилий закономерным образом распределено по регионам. В Западном регионе преобладают календарные фамилии - происходящие от имён православного календаря (Иванов, Васильев, Петров). В Центральном регионе - «звериные», включая и «птичьи», и фамилии от названий растений (Волков, Воробьев, Калинин). Для Восточного характерны как звериные, так и «приметные» фамилии (Смирнов, Румянцев, Белов). В Южном регионе чаще всего встречаются профессиональные фамилии (Попов, Гончаров, Ткачёв), а в Северном - календарные и иные (Булыгин, Лешуков, Чурсанов). И по самым частым фамилиям, и по частотам всех 14000 фамилий регионы средней полосы (Западный, Восточный, Центральный) оказались похожи. А Северный и Южный регионы отличаются и от средней полосы, и друг от друга.
Сравнение результатов. Было проведено сравнение 5 групп изученных признаков; учитывались три наиболее значимые компоненты по каждому из них (для мтДНК пока слишком мало изучено русских популяций, чтобы строить надёжные карты). Показана корреляция, сходство каждого группы с другими.
Почти все первые главные компоненты высоко коррелируют друг с другом. Особенно дружны оказались классические генетические маркёры и антропологические признаки, к ним присоединяется Y- хромосома и, с некоторым отрывом, дерматоглифика и фамилии. Конечно, между свидетельствами есть и различия, особенно на уровне менее значимых вторых и третьих компонент. Но в целом, пять разных типов признаков (несмотря на различную изученность, различный набор популяций, различные размеры выборок, разную информативность самих признаков и т. д.) обнаруживают единый строй генофонда. Поэтому можно утверждать, что выявлена реальная структура русского генофонда, которая пробилась сквозь все несовершенства методов исследований.
Выявленные закономерности показаны на рисунке 2. Мы видим, что все карты выявляют одно и то же - широтное направление изменчивости. Одни экстремумы расположены в северных областях, экстремумы противоположного знака занимают южные русские территории. В промежуточной зоне значения плавно изменяются.
Четыре карты незначительно различаются лишь двумя элементами: 1) относительным положением южного ядра (на разных картах оно смещается западнее или восточнее, или же выявляются сразу оба - юго-западное и юго-восточное ядра); 2) степенью выраженности северного "поморского" ядра.
О корреляциях. Общее сходство первых главных компонент по всем типам признаков можно измерить коэффициентом множественной корреляции. Его величина составила 0.9. Тогда коэффициент детерминации достигает 0.8. Это означает, что обобщённые карты самых различных признаков на 80% детерминированы некой единой темой структуры русского генофонда, и лишь на 20% допускают вариации на заданную тему. Эта единая тема - широтная изменчивость русского генофонда. Очень высокая гетерогенность русского генофонда как бы растянута по оси «север<=>юг», и, при движении в этом направлении, видны наибольшие различия между русскими популяциями.
Свидетельства археологии. «Ось Север — Юг сформировала геополитическую основу древнерусского государства» (Н. А. Макаров). Есть и другие важные свидетели, подтверждающие выявленные закономерности. Широтное деление Русской равнины (север, юг и серединка) согласуются с данными лингвистики о подразделении русского языка на северные, южные и переходные среднерусские говоры.
Археология ранее придерживалась иного мнения о Древней Руси - «Киево-центристской» концепции, где Северу отводилась роль второстепенной провинции. Однако накопление данных заставило пересмотреть это мнение: большинство археологов сейчас рассматривает становление Древней Руси как результат объединения двух равноправных государственных образований - Киева и Новгорода [Седов, 1999; Носов, 2002; Янин, 2004; Макаров, 2005]. Более того, Новгородские земли и Киевщина рассматриваются лишь как части двух обширных историко-культурных ареалов с различными направлениями внешних связей: Юг ориентировался на Причерноморье, а Север — на Балтику. Причём влияние Балтики простиралось на внутренние континентальные области Восточной Европы вплоть до Волго-Клязьменского междуречья, Белозерья и Северной Двины. Различия между Севером и Югом охватывали широчайшие пласты
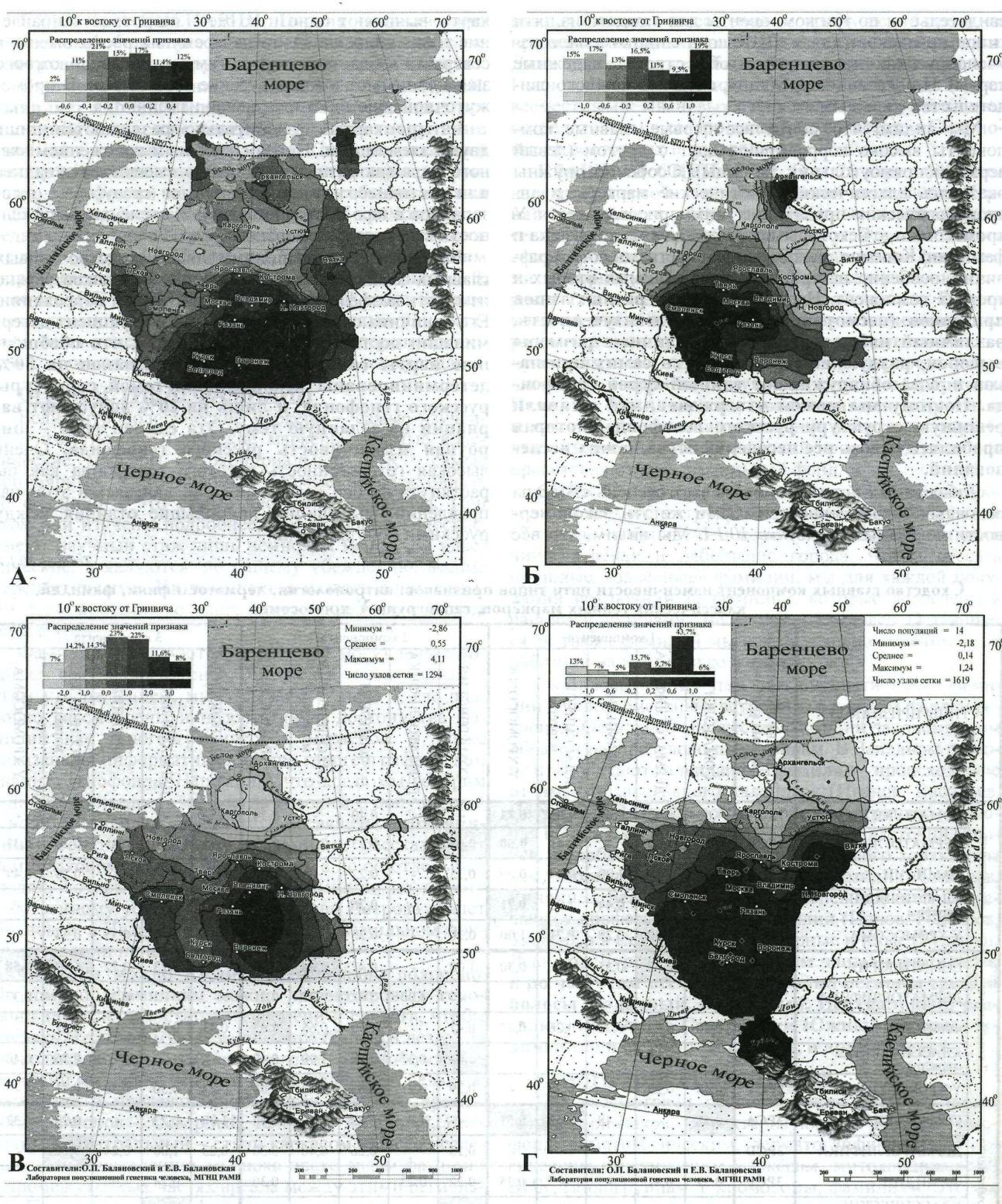
Рисунок 2- Особенности русского генофонда ( карты главных компонент изменчивости): A. Главный сценарий антропологии (соматология); Б. Главный сценарий дерматоглифики (кожные узоры); B. Главный сценарий классических маркёров генетики; Г. Главный сценарий антропологии маркёров Y хромосомы [2]
культуры: различия в керамике, градостроительстве, жилых постройках, сельскохозяйственных культурах, денежно-весовых системах, характере расселения. Причём мнение о бедности Севера по сравнению с цветущим Югом оказалось неверным - торговля пушниной позволяла большой части населения Севера жить в большем достатке и довольстве, чем на Юге, где богатство скапливалось у элиты. Своеобразной была и средняя полоса Русской равнины, в которой встречались культурные традиции Севера и Юга.
Но для генетики, конечно же, важно, насколько движение культуры было связано с миграцией самого населения, миграциями генов. И здесь тоже есть примечательные перемены в археологии. Прежние представления о массовых передвижениях восточных славян с юга на север в XI-XIII веках оказались преувеличенными. «Археологические материалы раскрывают многочисленные факты подвижек населения на север внутри регионов или через границы сопредельных областей, но не подтверждают картины массового отлива населения из южнорусских земель» [Макаров, 2005].
Это свидетельство не только вновь подтверждает два полюса русского генофонда и стрелку, указывающую на север (или на юг). Этот новый свидетель сообщает нам, что своеобразие Севера связано со своеобразием именно славянского мира. Важность этой информации трудно переоценить! Мы уже не раз встречались с тем, что своеобразие генофонда Русского Севера нельзя объяснить только мощным финно-угорским пластом. На него накладывался второй фактор - своеобразие северного славянского пласта генофонда. А особость культуры помогала сплавить все эти составные части в яркий и многоликий генофонд Русского Севера.
Русский генофонд в кругу соседей. Выявление широтной изменчивости в русском генофонде (ось «север<=>юг») тем удивительнее, что ожидать можно было лишь «противоположной» - долготной изменчивости (ось «запад<=»восток»). Ведь для генофонда Восточной Европы, который включает русский генофонд как составную часть, убедительно показана долготная изменчивость. А «исконный» ареал русского народа занимает основную часть Восточной Европы. Он сложился на основе восточнославянских, балтских и финно-угорских племён, а возможно также тюркоязычных и ираноязычных - едва ли не всех групп, населяющих восточную половину Европы. Поэтому основная закономерность генофонда Восточной Европы (долготная изменчивость) и должна была отразиться в русском генофонде, если он действительно складывался, как и Русское государство, «прирастанием», механическим включением в себя всё новых и новых групп населения. То, что выявлена иная, широтная изменчивость, указывает на принципиальный факт своеобразия русского генофонда. Это своеобразие состоит в том, что основное направление изменчивости русского генофонда не сводимо к исходной изменчивости тех субстратов, племён и народов, на основе которых он формировался. По-видимому, широтная изменчивость возникла или же усилилась в ходе собственной истории русского генофонда (т.е. в более поздние времена?).
Восточная Европа. Можно выделить несколько основных результатов изучения генофонда народов Восточной Европы.
Значительная межпопуляционная изменчивость. В Восточной Европе она намного больше, чем различия между популяциями в Западной Европе: этот результат подтвержден двумя свидетелями: и классическими маркёрами, и мтДНК.
Восточноевропейский генофонд сформирован главным образом народами индоевропейской и уральской языковых семей. Причём основная часть Восточной Европы похожа на результат длительного смешения этих двух генофондов. Что же касается народов третьей языковой семьи - алтайской, то их влияние меньше и ограничивается юго-восточными степными районами (этот результат также подтверждён двумя свидетелями: и аутосомными ДНК маркёрами, и классическими).
3) Средняя частота «азиатских» генов в Восточной Европе мала, и их распределение строго упорядочено - частота постепенно возрастает с запада на восток. Так что восточные славяне ещё полностью принадлежат европейской генетической провинции; на территории Волго-Уральского региона происходит постепенное увеличение частоты «азиатских» генов до значимых величин (10-30%), но не до 50%, то есть и этот регион преимущественно ещё «европейский»; лишь Западная Сибирь оказывается пограничной областью между западно-евразийским и восточно-евразийским генофондами (это данные по мтДНК, но и второй свидетель - Y хромосома - также не выявляет высоких частот «азиатских» гаплогрупп в Восточной Европе).
4) Первые главные компоненты (важнейшая закономерность) показывают долготную изменчивость Восточной Европы (классические маркёры, аутосомные ДНК маркёры, антропология, дерматоглифика, мтДНК).
Итак, важнейшая черта восточноевропейского генофонда - наличие в нём «западной» и «восточной» групп популяций. Генетические различия между ними и создают постепенный, плавный переход - долготную изменчивость.
История народов Восточной Европы богата переселениями, смешениями разнородных групп - от прихода с востока гуннов до экспансии на восток славян, многими другими миграциями, хуже известными нам из-за своей древности. Многие из них проходили по оси «запад<=>восток». Поэтому любое из этих событий (или все они вместе) могли сформировать долготную изменчивость.
Евразия. Надо сказать, что долготный тренд не ограничивается Восточной Европой. Он уходит далеко на восток и продолжается также на запад. Долготный тренд - основная закономерность генофонда всей Евразии. А долготный тренд Восточной Европы - лишь фрагмент, часть этого постепенного изменения, протянувшегося от Иберийского полуострова до Корейского. Эту глобальную закономерность евразийского генофонда особенно наглядно показывают карты, построенные для гаплогрупп митохондриальной ДНК. И мы опять видим постепенность этого тренда - огромные пространства заняты «промежуточными» генофондами. Напрашивается сопоставление между западно-евразийским генофондом и европеоидной расой, с одной стороны, и зоной преобладания восточно-евразийских гаплогрупп и монголоидной расой, с другой.
В геногеографии есть правило - чем тренд масштабнее, тем он древнее. И для глобального долготного тренда есть прямые доказательства его древности: проведённый картографический анализ археологических культур верхнего палеолита показал, что 26-16 тысяч лет назад уже существовали резкие различия между населением западных областей (Европа) и восточных областей (Сибирь).
Итак, долготный тренд - базовая, древняя закономерность Евразии, фрагмент этого тренда хорошо виден в Восточной Европе. Но в русском ареале, занимающем обширнейшую срединную часть Восточной Европы, на первый план вышла не эта фоновая, «подстилающая» долготная изменчивость, а иная, широтная закономерность, которая является собственной, характерной чертой русского генофонда.
Гетерогенность русского генофонда велика и заметна даже на картах «евразийского масштаба». Эта гетерогенность во многом описывается широтной изменчивостью, частично - множеством иных закономерностей. Достоверно известно: в масштабе Евразии, разделяемой в первом приближении на западные, восточные и промежуточные области, русский генофонд однозначно относится к западному стволу.
Азиаты ли мы? Стоит в любой аудитории упомянуть, что мы изучаем русский генофонд, как за этим всегда следует вопрос: насколько сильно его изменили татары? Иногда тот же вопрос звучит в иной формулировке: «сильно ли его изменили монголы?» И в обоих случаях имеют в виду одно и то же: не сходство с генофондом современных нам европеоидных татар-земледельцев, живущих в Приуралье, а генетическое сходство с теми ордами номадов, которые в XIII-XV веках шли из Центральной Азии и несли её чуждые гены в Европу. Этот вопрос нам задавали в самых разных аудиториях, не только в России, но даже в США. Похоже, самое распространённое представление о русском генофонде выражается фразой «поскреби русского и увидишь татарина». Сейчас мы попробуем честно ответить на этот многократно заданный вопрос.
Однако, каков вопрос, таков и ответ. Мы будем пользоваться (да простят нас профессионалы!) той же терминологией, что и задающие эти вопросы непрофессионалы. Иначе этот раздел превратился бы в лекцию о том «что иметь в виду под термином...», и заинтересованным лицам, не склонным к долгим научным уточнениям, это представилось бы способом уйти от прямого ответа. Нет, мы хотим дать максимально прямой и корректный ответ на непрофессионально заданный, но всех волнующий вопрос.
Чтобы не запутаться в словах, начнём с того, что современные татары и монголы - два разных народа, между генофондами которых очень мало общего. Поэтому мы будем говорить конкретно лишь о том, какие последствия для русского генофонда имело «татаро-монгольское иго», то есть завоевание Руси в ХIII веке и последовавшая зависимость русских княжеств от Золотой Орды и её наследников. Конкретнее: каков вклад в русский генофонд степных завоевателей ХIII века и нескольких последующих веков? Или ещё ближе к совопросникам века сего: стал ли (и если - да, то насколько) русский генофонд в результате этих завоеваний монголоидным - «с раскосыми и жадными очами» (А.Блок)? Естественно предполагать, что завоеватели оставляли своих потомков в местном населении, что завоевание сопровождалось и смешанными браками, и переселениями отдельных групп - наверное, ни у кого не вызывает сомнения, что при политическом подчинении одного народа другому происходит смешение их генофондов. Вопрос лишь, в какой степени.
«Завоеватели» происходили из степей Центральной Азии. Поэтому нам остаётся лишь определить, насколько русский генофонд стал похож на центрально-азиатский (а не на генофонд современных нам татар!).
Отделить генетический след «татаро-монгольского» ига от следа более древних и много более длительных взаимодействий генофондов запада и востока поможет сравнительная геногеография. Если, скажем, генофонд поляков (другого славянского народа, который монгольские завоевания затронули лишь отчасти, а трёхсотлетнее иго не затронуло вовсе) или же народов Севера Европы (куда никогда не доходили волны номадов) окажутся не похожими на центрально-азиатский генофонд, а русский генофонд, напротив, приближен к нему - то эту близость к центрально-азиатскому генофонду можно отчасти связывать с татаро-монгольским игом. Почему лишь отчасти? Потому что наличие такого сходства русского генофонда и Центральной Азии может быть результатом и более древних миграций, а вот если сходства не будет - это однозначно укажет на отсутствие «татарского следа». Всё это - правдоподобная модель, а реальность, конечно, сложнее. Но популяционная генетика всегда пользуется относительно простыми моделями, чтобы получить чёткий количественный ответ, а потом уже двигаться дальше - с помощью уточнённых моделей.
Итак, вопрос о «генетическом наследии татаро-монгольского ига» мы формулируем так: насколько русский генофонд похож на генофонд Центральной Азии на фоне отсутствия такого сходства у народов, не бывших «под игом».
Данные по митохондриальной ДНК. По этому типу маркёров народы Центральной Азии и Европы различаются замечательно чётко: в Центральной Азии почти всё население - более 90% - обладает восточно-евразийскими гаплогруппами, тогда как в Европе почти всё население - более 95% - обладает иными, западно-евразийскими гаплогруппами. Поэтому доля восточно-евразийских гаплогрупп в русском генофонде прямо покажет: есть ли заметный вклад центрально-азиатского населения в русский генофонд или его нет?
Сразу же договоримся, что считать эффектами смешения и «заметным вкладом» - 0.5%, 5% или 50% восточно-евразийских гаплогрупп в русском генофонде? Учтём, что подразделение на западно - и восточно-евразийские гаплогруппы не абсолютно. Итак, если в Европе (включающей всё население до восточных склонов Урала) примерно 5% генофонда «не западные», а в Центральной Азии примерно 10% генофонда «не восточные», то наличие 15-20% восточно-евразийских гаплогрупп могло бы чётко маркировать присутствие «монголоидного» центрально-азиатского пласта в русском генофонде. 20% восточно-евразийских гаплогрупп означало бы приток 18% «монголоидных» генов в русский генофонд, а 15% восточно-евразийских гаплогрупп - приток 13% центрально-азиатских генов. Если кто-то желает увидеть ещё менее значительные и почти неощутимые влияния, то чтобы распознать их, ему надо провести анализ очень объёмных выборок - иначе результаты будут недостоверными.
Не азиаты мы... увы... Однако доля восточно-евразийских гаплогрупп в русском населении составляет лишь 2%. То есть очень малую величину. Почти столь же малую, что и в генофонде поляков (1,5%) или севера Европы, где уж точно никаких «татаро-монгольских вкладов» не было (у норвежцев 0,6%, у исландцев 0,7%, у карел 4,8% и т. д.). Средняя «фоновая» частота восточно-евразийских гаплогрупп в Европе равна 3,6%. То есть в русском генофонде (2,0%) она даже меньше, чем «средняя по Европе», поэтому монголоидный компонент у русских оказывается не просто нулевым, но даже с отрицательным знаком. Итак, мы не видим последствий монгольского нашествия в русском генофонде - или тогда должны видеть эти последствия и в польском, и в норвежском, и в карельском, и в других генофондах Европы.
Не ошиблись ли мы? Давайте проверим. Все использованные данные, то есть частоты гаплогрупп мтДНК, получены по большим выборкам, самыми разными исследователями, и поэтому вполне достоверны. Но может быть, армии Чингисхана и Батыя состояли не столько из населения Центральной Азии, сколько из степных народов Южной Сибири? Но и в Южной Сибири восточно-евразийские гаплогруппы составляют пусть не полные 90%, а только 60-80%, но это всё равно несопоставимо больше, чем 2%! Пожалуй, иной читатель - не желающий расставаться с романтической картинкой «монгольского следа» - предпочтёт предположить, что монгольские армии состояли из европеоидных воинов! Но мы-то проверяем мнение о «нас - азиатах», а такой читатель в пылу полемики начинает защищать уже совсем иной миф - что «татаро-монголы» ничем от населения Европы генетически не отличались. Но такой читатель забывает, что тогда снимается и сама проблема - если генетически Золотая Орда и её наследники были, по его мнению, европейцами, то это уже становится внутренним делом Европы: перемещения народов в её пределах. А ведь мы решаем проблему влияния Азии на генофонд Европы.
Может быть ещё одно возражение: мтДНК наследуется по материнской линии, а «генетический вклад завоевателей» ожидается скорее по отцовской. Это возражение отметать нельзя, поэтому рассмотрим данные по Y хромосоме («мужская», отцовская линия наследования).
Данные поY- хромосоме. Результаты изучения гаплогрупп Y- хромосомы также не показывают значительной доли «степных монголоидных» генов в русском генофонде. Среди восьми гаплогрупп, преобладающих в русском генофонде, есть «восточноевропейская» R1a, «скандинавская» IIа, «балканская» lib, «западноевропейская» R1b, «ближневосточная» J2, «африкано-средиземноморская» Е3b, «западносибирская» N2, «восточноевропейско-сибирская» N3 - но нет «степных», «центральноазиатских» гаплогрупп. Правда, Rla встречается и в степях Сибири - но реже, чем у русских. Гаплогруппа N3, частота которой в Сибири выше, чем у русских, остаётся единственным кандидатом на «степное» происхождение (хотя эта гаплогруппа скорее «таёжная») - но, увы, она встречена лишь в северных русских популяциях, а генетическое влияние степного населения должно было бы проявиться в южных районах исторического русского ареала. В любом случае - N3 распространена по всему северу Восточной Европы, и у эстонцев, финнов, латышей встречается куда чаще, чем у русских, так что монгольские армии тут явно ни при чём.
Антропологические данные. Любопытны данные по эпикантусу - типичнейшему признаку монголоидной расы, особенно характерному как раз для степного населения Центральной Азии. Антропологическое исследование, проведённое на огромной выборке русских (несколько десятков тысяч человек) практически не выявило случаев выраженного эпикантуса [Хрисанфова, Перевозчиков, 1999].
Итак, какой бы признак мы ни взяли, мы видим, что русские - типичные европейцы, а азиатское завоевание оставило след в русской истории, но не в русском генофонде.
Так откуда же взялось столь распространённое мнение, что генетически «мы - азиаты», или, по крайней мере, промежуточное звено между Европой и Азией? Увы, это мнение распространено и в научной среде. Мы уже упоминали про один - но характерный! - случай, когда по какому-то показателю русские оказались промежуточны между французами и китайцами, и Учёный совет, состоящий из прекрасных учёных, счел это совершенно естественным. Мнение о промежуточности русского генофонда столь широко распространено - но ему нет серьёзных научных подтверждений. Одни лишь серьёзные научные опровержения.
Не русскому генофонду выпала роль «буферной зоны» между западом и востоком, не он стал местом их встречи - эта роль досталась иным народам, живущим на восток от Урала, в Западной Сибири. А русский генофонд остался в строю народов европеоидных, западно-евразийских.
При этом русский генофонд является самым восточным из типично европейских; отдельные азиатские влияния прослеживаются в русском генофонде несколько больше, чем у его западных соседей; уровень гетерогенности европейских генофондов мал, сибирских - велик, а восточноевропейские генофонды, в том числе русский, промежуточны (но не по генетическому составу, а по уровню гетерогенности!) между западноевропейскими и сибирскими; структура же современного татарского генофонда и его взаимодействие с русским куда сложнее, чем может показаться на первый взгляд, и заслуживает отдельной монографии.
Но базовый, главный вывод, который следует из проведённого изучения русского генофонда - это практически полное отсутствие в нём монголоидного вклада. Даже исследуя исключения, никогда нельзя забывать это правило.
Интересна гипотеза Е.В. и О.П. Балановских. Последствия «татаро-монгольского ига» для русского генофонда, вероятно, есть, но заключаются совсем не в том, как это представляется обычно. Не во вкладе «центрально-азиатских генов», а в изменении миграций собственно русского населения. То есть в изменении направления и интенсивности потоков генов, которые, в свою очередь, сказались на русском генофонде, в какой-то степени перестроив его структуру. Возможно, лишь в небольшой степени и только для восточной части русского ареала.
Известно, что монгольское завоевание застало Киевскую Русь в период феодальной раздробленности. Часть древнерусских княжеств попала в зависимость от Золотой Орды, и на этой территории впоследствии сформировался русский народ. Другая часть древнерусских княжеств вошла в состав Литовского (позже - польско-литовского) государства, и образовала украинский и белорусский народы. Разделение древнерусской народности на разные субэтносы - это ли не значимые последствия монгольского завоевания!
Восточные славяне - в течение тысячелетия, с беспримерной энергией - колонизировали огромные пространства, от Днепра и до Волги, до Енисея, до Юкона. Единственная пауза в этом многовековом процессе - период ига, когда восточное направление было закрыто Золотой Ордой и её наследниками. В эти столетия проходила так называемая внутренняя колонизация русских земель, и не на эти ли века вынужденного сидения в своих границах пришлось само формирование русского народа?
В период ига постоянно происходили войны с татарами в приволжских землях. (Имеется в виду среднее течение Волги примерно от Костромы до Казани, и соответствующие притоки Волги - Унжу, Оку, Ветлугу, Суру). Войны вызывали переселения, истребление селений, пленения... Не приходится сомневаться, что все эти события влияли на миграции и демографическую структуру русского населения. Но был ли при этом генетический вклад «татарского компонента»?
Русское население по реке Унже было изучено по митохондриальной ДНК. Суммарная частота восточно-евразийских гаплогрупп составила 4%, при среднем уровне в русских популяциях 1-2%. У современных татар частота восточно-евразийских гаплогрупп составляет около 10%. Такие невысокие частоты и недостаточные объёмы обеих выборок не допускают точных вычислений. Но даже если считать двухпроцентное повышение частоты реальностью, всё равно оно слишком мало. А главное, повышение частоты восточно-евразийских гаплогрупп к востоку это, как говорилось, общий евразийский тренд, древность которого измеряется тысячелетиями. И чуть повышенная частота у русских Унжи, скорее всего, вызвана смешением не с татарами, а с финно-угорским племенем меря (родственным современным мари), жившем на Унже до прихода славян.
Ещё раз подчеркнём: влияние татарских государств - это совсем не тот вклад «центрально-азиатского генофонда», возможность которого мы обсуждали чуть выше. Совсем не тот хотя бы потому, что генофонд как современных, так и средневековых татар составлен из многих компонентов, и «азиатский» степной компонент, возможно, не главный в нём, как мы видим и из невысокой частоты восточно-евразийских гаплогрупп у татар. Вся сложность задачи реконструкции истории на основе современного татарского генофонда как раз в том, что «поскреби татарина и увидишь финно-угра» - тот же самый финно-угорский пласт, лежащий в основе и русского генофонда на востоке его ареала.
Таким образом, мини-исследование Унженского края не обнаружило особого вклада «татарского компонента». Вероятно, влияние татарских государств заключалось скорее в вызванном ими оттоке населения, изменении структуры, закономерности миграций - то есть в демографических, а не генетических изменениях. Это совершенно разные процессы, и их нельзя путать: одно дело - миграция «местных русских генов» в пределах русского же ареала (реально имевшая место в связи с русско-татарскими войнами), а другое дело - прямой генетический вклад «пришлых татарских генов» (небольшой или вообще отсутствующий).
Итак, анализ русского генофонда однозначно показал практически полное отсутствие вклада монголоидного населения (азиатских степей). Ограниченное взаимодействие с татарским генофондом (европейских степей) возможно, имело место. Но не заметно, чтобы интенсивность смешения русского населения с татарским превышала обычный уровень смешений в зоне контакта двух народов. Нелишне напомнить, что русский генофонд контактирует со многими народами - Северного Кавказа, с украинцами, белорусами, латышами, эстонцами, финнами, карелами, коми, удмуртами, марийцами, мордвой и - в этом общем ряду - и с татарами.