

2. Формирование градиентов в яйцеклетке
Как гены контролируют формирование градиентов, а следовательно, и план строения будущего организма в ходе оогенеза? Естественно, что наиболее удобным объектом исследования этого контроля является знаменитая плодовая мушка дрозофила в силу ее хорошей генетической изученности.
Яйцеклетка дрозофилы развивается из общей с трофоцитами клетки-предшественника. Эти клетки-предшественники выделяются очень рано, после того как ядра будущих половых клеток попадают в ту область полярной плазмы, в образовании которой важную роль играет ген oscar.
Каждая такая клетка претерпевает 4 деления, так что возникает 16 клеток, одна из которых будет половой. Остальные клетки становятся питающими (трофоцитами) и соединены с яйцеклеткой цитоплазматическими мостиками, по которым в нее поступают различные вещества, принимающие участие в формировании градиентов. ДНК хромосом трофических клеток претерпевают многократное деление (так называемая политенизация), что существенно активизирует их функционирование в процессе обслуживания яйцеклетки.
Из материнской ткани образуется также около 1000 мелких фолликулярных клеток, окружающих будущий ооцит с трофическими клетками.
Будущий передний конец эмбриона располагается в области яйцеклетки, прилежащей к трофоцитам, а более выпуклая поверхность яйца становится брюшной частью эмбриона.
Далее начинается активное функционирование материнских генов в питающих клетках. Можно выделить три системы генов, особенно важных для формирования градиентов. Первая система генов обеспечивает формирование анимально-вегетативного (головно-хвостового) градиента (рис. 2). Вторая — дорзо-вентрального (спино-брюшного) градиента. Третья — синтез продуктов, необходимых для формирования специфических головных и хвостовых структур.
Наиболее изучена первая система генов. Среди них главным является ген bicoid, содержащий так называемый гомеобокс, специфическую, консервативную последовательность ДНК из 180 нуклеотидных пар. В случае его мутации нарушается развитие головного конца дрозофилы. У эмбрионов — носителей мутации по этому гену — задняя часть развивается нормально, но нарушено развитие передних брюшных сегментов, а вместо головы и груди развиваются структуры, свойственные заднему концу.
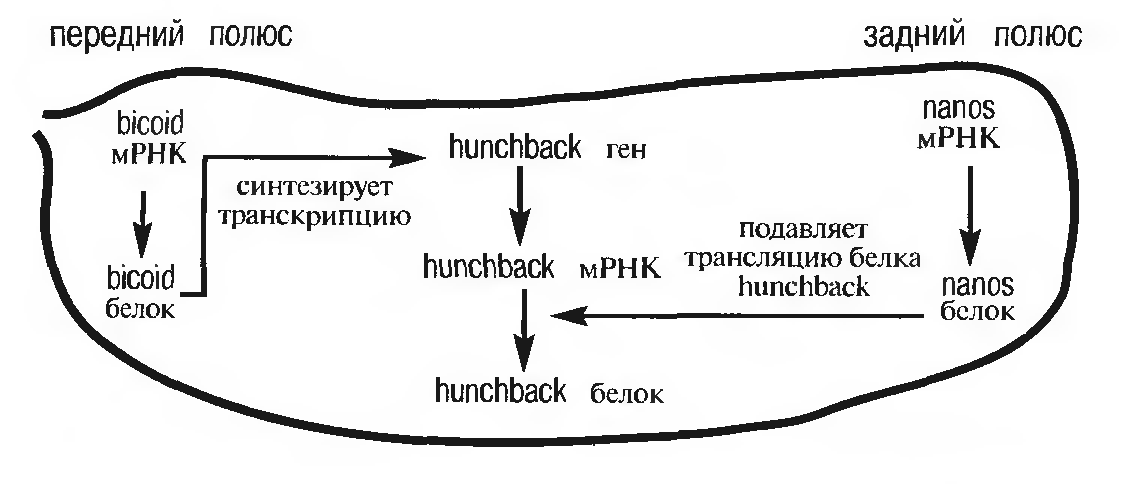
Рисунок 2 - Схема взаимодействия генов, отвечающих за формирование передне-заднего градиента яйца [1]
Если методами молекулярной генетики исследовать локализацию соответствующей матричной РНК, то можно проследить ее транспорт из питающих клеток в передний полюс развивающегося ооцита. В результате формируется четко выраженный анимально-вегетативный градиент распределения продукта этого гена. Белки, управляющие программой развития зародыша, принято называть морфогенами.
С другой стороны, питающие клетки, окружающие задний полюс яйцеклетки, «поставляют» в нее РНК, синтезированную геном nanos. У мутантов nanos нарушается развитие заднего конца зародыша. Если nanos РНК инъецировать в передний конец эмбриона, она может индуцировать формирование в головном конце различных структур, свойственных заднему полюсу. Белок nanos синтезируется в области заднего полюса и затем транспортируется в область брюшных сегментов.
В формировании плана строения организма на самых ранних этапах созревания яйцеклетки принимает участие еще один очень важный ген, который активно функционирует не только в материнском, но и в зиготическом геноме (т. е. в геноме самой оплодотворенной яйцеклетки). Это ген hunchback. Он активируется белком bicoid, а потому его продукт накапливается, как и bicoid, в передней половине зародыша и подавляет гены, активные в брюшных сегментах, так что в зоне его распределения формируются головные и грудные структуры.
В норме белок nanos тормозит синтез белка материнской РНК hunchback. Именно это и является основной его функцией. Если имеют место мутации, нарушающие функционирование как nanos, так и hunchback, то развивается нормальный организм.
Отсюда следует, что продукт гена hunchback «мешает» развитию хвостового конца, а блокирующий эффект со стороны гена nanos как бы устраняет чинимые им помехи (рисунок 3).
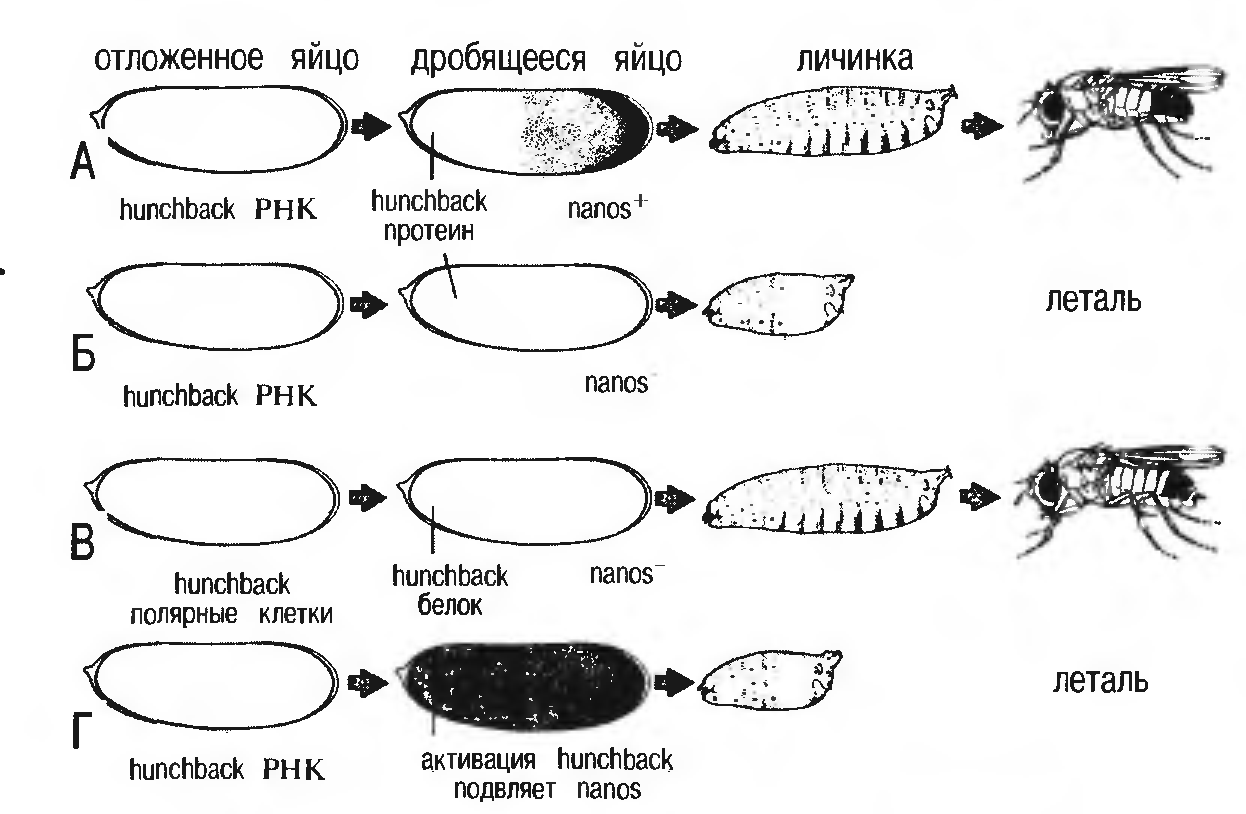
Рисунок 3 - Схема регуляции развития зародыша дрозофилы с участием генов hunchback и nanos [1]
A. Вот что происходит в норме. В отложенном яйце матричная РНК hunchback распределена равномерно по всему объему. Однако белок hunchback (как запасенный до оплодотворения, так и синтезированный в яйце после оплодотворения) в заднем полюсе отсутствует, поскольку функция его матричной РНК блокирована белковым продуктом гена nanos, накапливающимся в хвостовом отделе. Hunchback не дает развиваться хвостовым структурам в переднем отделе эмбриона, а в задних отделах ген nanos блокирует его синтез, и хвостовые структуры развиваются, как и положено в норме.
Б. А вот что случается, если ген nanos мутировал и потерял способность «работать». Поскольку его продукт отсутствует, продукт гена hunchback распространяется по всему яйцу и мешает формированию хвостовых структур. Развитие нарушается, и на стадии личинки муха погибает.
B. Однако, если наряду с отсутствием продукта гена nanos в отложенной яйцеклетке в результате соответствующей мутации подавлен синтез продукта гена hunchback, ничто не мешает формированию хвостовых структур.
Головные структуры также формируются, поскольку в оплодотворенной яйцеклетке продукт гена hunchback накапливается только в передней (головной) области эмбриона. Налицо, таким образом, все условия для нормального развития.
Г. Если с помощью специальных методов заставить ген hunchback работать с повышенной активностью, то белка nanos не хватит для того, чтобы целиком блокировать синтез его белкового продукта, даже если ген nanos нормален. Тогда белок hunchback накапливается в достаточно большом количестве, чтобы помешать формированию хвостовых структур.
Вторая и третья системы генов контролируют формирование спино-брюшного градиента и работают по тому же самому принципу, как и первая, только участвуют в этой «работе» другие гены.
Итак, неоднородность цитоплазмы созревающей яйцеклетки и формирование полярных градиентов, химически преформирующих план строения будущего организма, реализуется на основе взаимодействия трех систем генов и при участии питающих клеток материнского организма, окружающих ооцит.