

Тема № 8. Генетика индивидуального развития
Краткое содержание:
1. Ооплазматическая сегрегация и полярная плазма
2. Формирование градиентов в яйцеклетке
3. Гены сегментации
4. Гомеозисные гены, их роль в развитии
5. Гипотеза Э. Льюиса о механизме функционирования гомеозисных генов 6. Гомеобокс и гомеодомен. Принцип коллинеарности
7. Гены — господа и гены — рабы. Опыты Вальтера Геринга
Одна из ключевых проблем современной биологии — проблема индивидуального развития. Этой проблемой занимается особый раздел генетики - генетика развития, которая оформилась на стыке, по крайней мере, трех научных дисциплин — генетики, эмбриологии и молекулярной биологии. Именно комбинация этих методических подходов позволяет проследить путь от гена к формированию зародыша и, в конце концов, - к признаку у взрослого организма.
Совсем недавно были открыты новые гены и генные каскады, знание которых преобразило наши представления о законах индивидуального развития.
1. Ооплазматическая сегрегация и полярная плазма
Какой момент можно считать началом индивидуального развития? Первое деление дробления? Оплодотворение яйцеклетки? Томас Морган справедливо заметил, что за точку отсчета надлежит принимать еще более ранний период времени, а именно — созревание яйцеклетки. Он писал, что неправильно считать ее недифференцированной системой, что на самом деле она является самой высокоспециализированной клеткой в организме, поскольку именно в ходе ее созревания закладывается план будущего строения организма.
Этот тезис Моргана ставит этические проблемы при работе с бластоцистами (самыми ранними эмбрионами) человека с целью получения из них «запчастей» для трансплантации. Согласно этому тезису, живая бластоциста является полноценным зародышем человека, и экспериментальные манипуляции с ней означают убийство.
И действительно, развитие яйцеклетки, по сути дела, представляет собой последовательное формирование неоднородности ее цитоплазмы, в результате чего осуществляется так называемая ооплазматическая сегрегация. В этот период функционируют почти все гены, так что в яйцеклетке содержится набор самых разнообразных матричных РНК (мРНК), ответственных за синтез белков, многие из которых станут нужными лишь на поздних стадиях развития. Ядро развивающейся оплодотворенной яйцеклетки функционирует, следовательно, как бы с опережением, работая не только на настоящее, но и на будущее.
В ходе ооплазматической сегрегации формируются те особенности отдельных участков цитоплазмы, которые как бы намечают, «преформируют» на химическом уровне план строения будущего организма. В основу формирования этого плана лежит образование так называемых полярных градиентов распределения биологически активных веществ.
Полярный градиент — это неравномерное распределение вещества вдоль оси зародыша с постепенным падением его концентрации от одного полюса к другому.
В яйцеклетке имеется несколько таких градиентов, в частности, передне-задний (анимально-вегетативный) градиент, который проявляется в постепенном падении концентрации РНК и белков, уменьшению активности синтеза РНК и белков в направлении от анимального (головного) к вегетативному (хвостовому) полюсу. В вегетативной области яйца, напротив, сосредоточены инертные запасные питательные вещества.
Иными словами, яйцеклетка — отнюдь не гомогенное образование, но гетерогенная, химически преформированная (предобразованная), высокоспециализированная система. Значение этой преформации может быть выявлено экспериментально. Например, бельгийский цитолог и эмбриолог Жан Браше с помощью использования разных режимов центрифугирования нарушал градиентное распределение вещества цитоплазмы двумя способами: 1) разрушал систему градиентов, вызывая равномерное распределение РНК, белков и других веществ по цитоплазме; и 2) расчленял единый передне-задний градиент на два самостоятельных градиента.
Эти нарушения по-разному сказывались на судьбе развивающегося зародыша. В первом случае развитие останавливалось на самых ранних стадиях, и зародыш погибал. Во втором случае возникал двухголовый зародыш.
В чем же дело? По-видимому, в том, что неравномерное распределение РНК ведет к специфическому синтезу соответствующих белков в разных частях развивающегося зародыша. Уже на ранних стадиях его развития, в ходе дробления бластомеры (клетки, образующиеся при дроблении) приобретают определенные свойства, отличающие их друг от друга как в физиологическом, так и в молекулярном отношении.
У некоторых организмов, например у асцидий, неоднородность цитоплазмы яйцеклетки можно заметить по разнице в окраске разных ее частей. Достаточно рано проявляется у них и неоднородность в распределении некоторых белков.
Иногда специфически выделяется та или иная часть цитоплазмы, как например, полярная плазма яиц некоторых насекомых, включая дрозофилу, формирующаяся на заднем полюсе яйца, богатая РНК и характеризующаяся четко выраженной зернистой структурой. Клеточные ядра, которые попадают в эту цитоплазму, дают начало половым клеткам.
Если область полярной плазмы облучить ультрафиолетом, то развившиеся из таких локально облученных яиц зародыши будут стерильны, поскольку оказываются лишенными половых клеток. Микроинъекции полярной плазмы в различные участки развивающегося раннего зародыша вызывают образование половых клеток в необычном месте.
Возникает вопрос, чем вызвана ооплазматическая сегрегация, от каких факторов зависит формирование градиентов? Это связано с особым положением ооцита (оплодотворенных яйцеклеток) в материнском организме. Дело в том, что, начиная с самых ранних стадий формирования яйцеклетки, ее полюса находятся в неравном положении относительно окружающих их трофических (питающих) клеток. Всегда один из полюсов ооцита окружен большим количеством питающих клеток, снабжающих его через специальные канальцы различными веществами — РНК, белками, а у некоторых жуков даже и митохондриями (рис. 1).
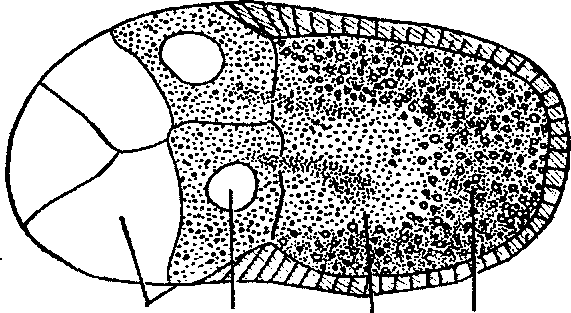
1 2 3 4
Рисунок 1 - «Миграция» клеточных органов — митохондрий из питающих клеток
в яйцеклетку: 1 — питающие клетки, 2 — ядро, 3 — митохондрии, 4 — ооцит [1]
Таким образом, поставленный в более благоприятное положение относительно питающих клеток полюс ооцита, становится впоследствии, анимальным (головным). Ясно, что формирование анимально-вегетативного градиента зависит от влияния материнского организма.