

3. Гены сегментации
В реализации плана строения организма ключевое значение имеет его сегментация, разделение на головной, грудной, брюшной отделы и их производные.
Этот процесс является универсальным в животном мире и характеризуется двумя основными признаками — количеством сегментов и их качеством. Соответственно различают две группы генов, ответственных за развитие этих признаков: сегрегационные и гомеозисные.
Сегрегационных генов, определяющих число сегментов у плодовой мушки дрозофилы, известно более двух десятков, их мутации вызывают нарушения в развитии передне-задней полярности сегментов, в результате чего происходит их слияние, уменьшение количества и образование нежизнеспособных уродов. За открытие и изучение этих генов американский генетик Эдвард Льюис и немецкие генетики Кристина Нюссляйн-Вольхардт и Эрик Вишхаус были удостоены Нобелевской премии.
Различают несколько групп сегрегационных генов. Это гены «материнского эффекта», которые контролируют формирование градиентов в ходе развития (о них шла речь выше), gap гены, pair-rule гены, гены сегментарной полярности, последовательно осуществляющие сегментацию зародыша и подготавливающие почву для функционирования гомеозисных генов. Следует особо отметить, что гены сегментации начинают функционировать в тот период, когда морфологически и следов-то сегментации нет, задолго до того, как сегментация морфологически оформится, т. е. они также работают с опережением, «на будущее».
Сегрегационные гены последовательно активируются в процессе индивидуального развития (рис. 4). В первую очередь активируются так называемые gap гены (от англ. gap — разрыв, щель). Их функция — синтез матричной РНК — стимулируется продуктами генов материнского эффекта, формирующими градиенты в ходе созревания яйцеклетки.
В результате данного процесса зародыш подразделяется на несколько пространственных отделов со своей химической спецификой. На фоне специфического распределения продуктов gap генов, под влиянием этих продуктов активируются pair-rule гены, которые «дробят» зародыш на еще более мелкие повторяющиеся части (рис. 5).
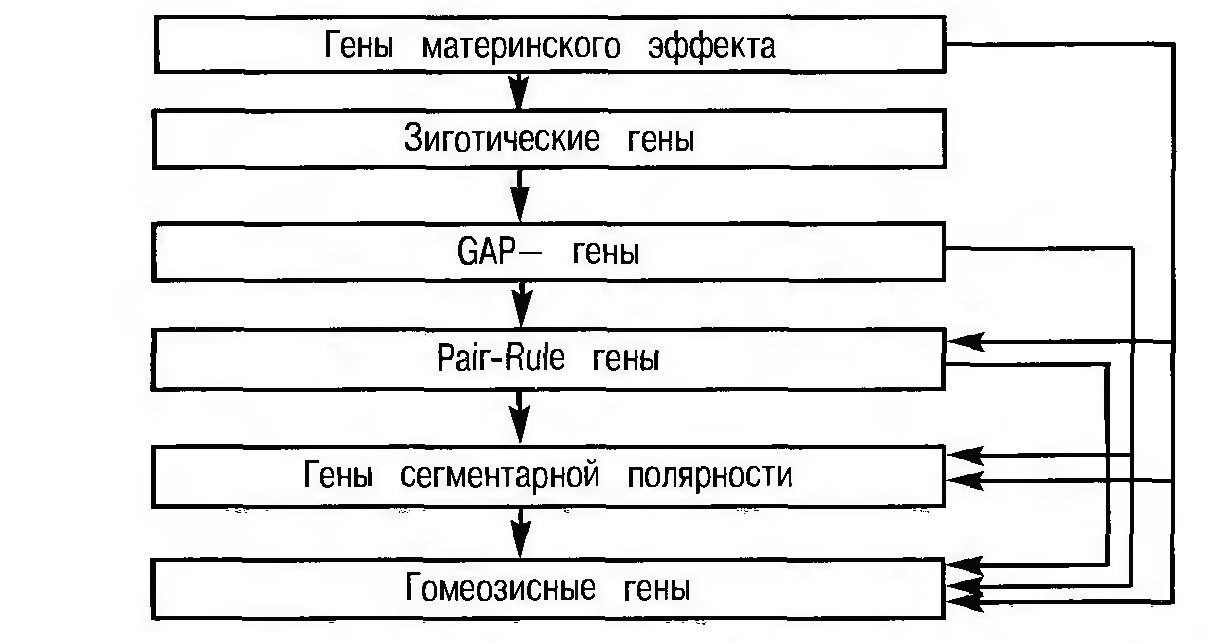
Рисунок 4 - Схема взаимодействия генов сегментации у дрозофилы [1]
Все сегрегационные гены, последовательно активируемые в ходе развития эмбриона дрозофилы, взаимодействуют друг с другом и оказывают друг на друга взаимные влияния через кодируемые ими продукты. Активация сегрегационных генов подготавливает «почву» для функционирования ключевой системы генов, обеспечивающих качественную спецификацию сегментов, — системы гомеозисных генов.
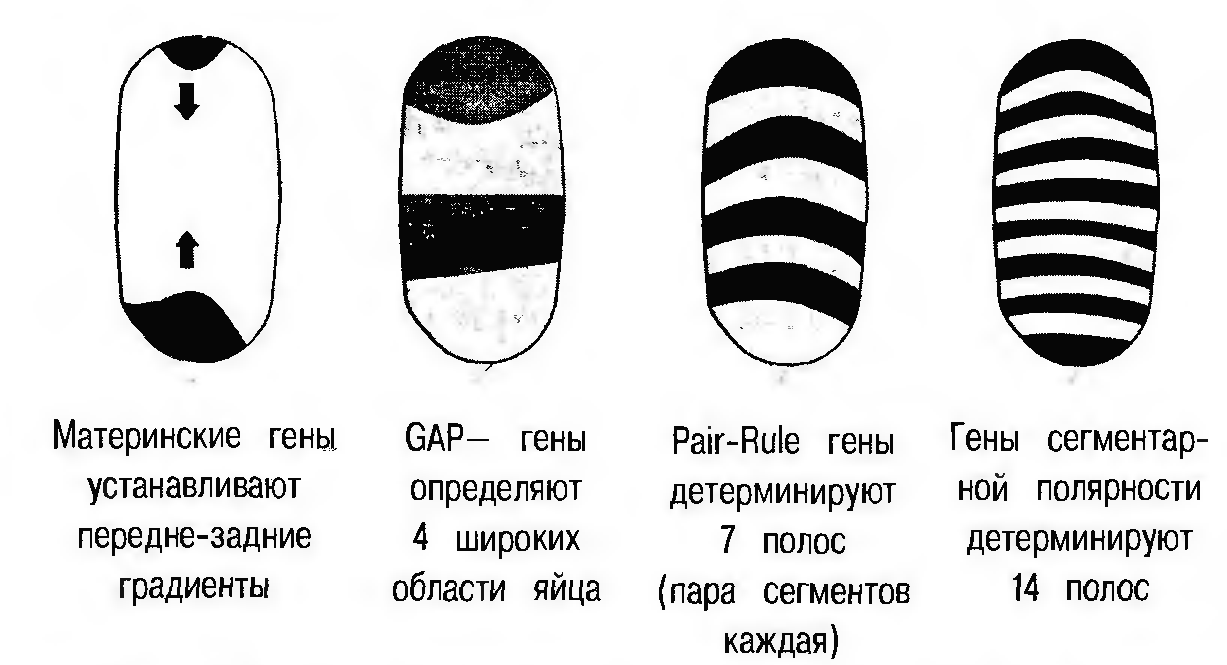
Рисунок 5 - Последовательное проявление генов сегментации. Различным образом заштрихованные полоски отражают последовательную гетерогенизацию развивающегося эмбриона дрозофилы [1]
4. Гомеозисные гены, их роль в развитии
Название этой группы генов происходит от термина «гомеозис», который ввел в 1894 г. один из классиков генетики Уильям Бэтсон. Под гомеозисом он понимал превращение одной части тела в другую. Гомеозисные гены, следовательно, не представляют собой нечто самостоятельное, но являются частью специфической системы генов, контролирующих сегментацию тела насекомых, в частности, дрозофилы, и других организмов. Примером гомеозисных мутаций является превращение антенны или аристы дрозофилы в ногу (рис. 6).
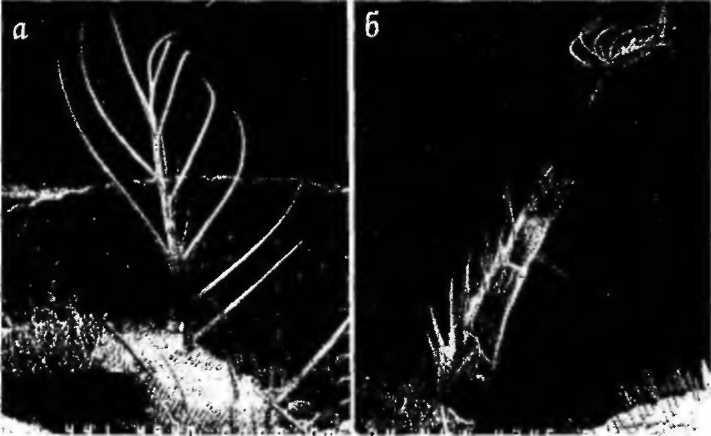
Рисунок 6 - Пример гомеозисной мутации. Превращение осязательного органа дрозофилы — аристы (слева) в ногу (справа) [1]
Весьма курьезный случай отмечен в хирургической практике: на голове пациента вырос половой член, который хирургам пришлось удалять, и который при гистологическом анализе обнаружил наличие всех свойственных нормальному половому члену структур. Возможно, это был вариант гомеозисной мутации у человека.
Гомеозисные гены, которых у дрозофилы описано около полусотни, как уже отмечалось, контролируют качественные особенности сегментов и в свою очередь подразделяются на два комплекса: Antennapedia-Complex (ANT-C) и Bithorax-Complex (ВХ-С).
Гены, принадлежащие к ANT-C, контролируют развитие головных сегментов, при утрате функции гена Antp грудные сегменты приобретают свойства головных сегментов, что проявляется в образовании головных структур в грудной области. Таким образом, от определенных генов зависит выбор программы развития, определяется путь, по которому клетка будет «двигаться» дальше, во что превращаться — в кожную, в нервную, в головную или брюшную клетку.
Гены комплекса ВХ-С ответственны за развитие грудных и брюшных сегментов. Рассмотрим действие гомеозисных генов на примере ВХ-С комплекса (рисунок 7).
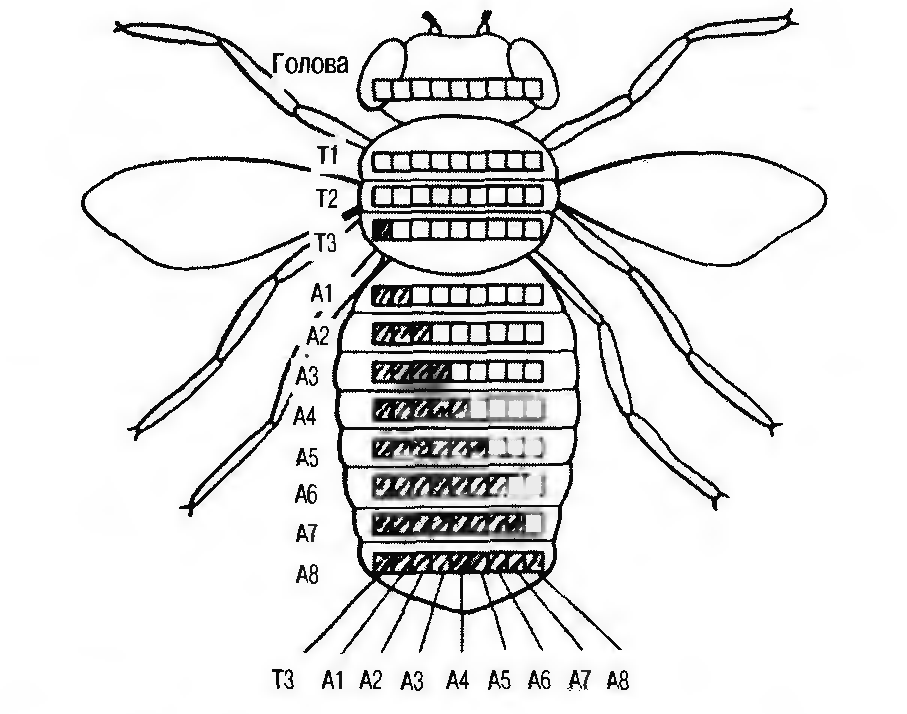
Рисунок 7 - Схема генетической регуляции процесса сегментации тела дрозофилы. В сегменте Т2 активен ген, мутация которого вызывает превращение Т3 в Т2. В каждом последующем сегменте активируется дополнительный ген. В последнем, 8-м сегменте работают все гены. От набора работающих генов зависят свойства сегментов [1]
Этот комплекс состоит из трех «отделов», которые называются: Ultrabithorax (Ubx), ответственный за развитие грудных сегментов (Т1-Т3), а также Abdomen-A и Abdomen-B, контролирующие дифференцировку брюшных сегментов (А1-А8). Все они построены и функционируют по единому принципу.
Так, например, гены, входящие в состав области Ubx, характеризуются двумя основными особенностями. Во-первых, они собраны в единый блок, так называемый кластер в небольшом участке 3-й хромосомы. В этот кластер входит пять генов, последовательно контролирующих развитие того или иного сегмента. Инактивация каждого такого гена вызывает превращение соответствующего сегмента в «двойника» впереди лежащего сегмента. Вторая особенность — коллинеарность в расположении генов и контролируемых ими признаков. Она заключается в том, что положение генов в комплексе ВХ-С соответствует последовательности контролируемых ими органов.