

5. Гипотеза э. Льюиса о механизме функционирования гомеозисных генов
Первооткрыватель гомеозисных генов дрозофилы Эдвард Льюис (США) сразу оценил их ключевое значение, как в индивидуальном, так и в историческом развитии и выдвинул предположение о механизме их функционирования.
«Нормальные» (немутировавшие) гомеозисные гены продуцируют вещества, дающие морфогенетический эффект (морфогенез — формообразование), тогда как мутанты не способны синтезировать их. Например, продукт нормального гена bxd подавляет потенциальное развитие 1-го брюшного сегмента по типу заднегруди, а продукт мутанта не способен это сделать.
Естественно, утрата способности синтезировать вещества, которые блокируют потенциальное формообразование в определенном направлении, ведет к осуществлению подавленных формообразовательных тенденций в другом направлении.
Предположение Льюиса несет определенный эволюционный смысл. Действительно, мутация bxd, которая приводит к неспособности синтезировать белок BXD, может быть аналогична тем, что обусловили в историческом прошлом превращение многоножек в предков современных шестиногих насекомых, т. е. у предков на брюшных сегментах формировались грудные структуры (ноги, крылья), но мутация типа bxd — bxct подавила эту способность.
Следовательно, зачатки, формирующие сегменты, первоначально обладали широким потенциалом развития, который был сужен последовательным рядом мутаций. Этот способ исторического развития является хорошим примером единства индивидуального и исторического развития. В некоторых случаях гомеозиса (превращения одной части тела в другую) примерно ясна адаптационная и эволюционная значимость преобразований.
Таким образом, смысл гомеозисных мутаций — изменение плана индивидуального развития с возможным филогенетическим выходом.
По результатам молекулярно-генетических исследований комплекса ВХ-С были установлены следующие особенности строения и функционирования гомеозисных генов:
1. Подавляющее большинство мутаций ВХ-С связано со вставками или вырезанием мобильных генетических элементов.
2. При анализе участка ДНК, содержащего Ubx, было выявлено его экзон—интронное строение, т. е. чередование кодирующих и некодирующих участков в ДНК.
3. Время первого появления транскриптов ВХ-С очень раннее — уже на 2—4-м часах после оплодотворения яйца. Таким образом, гены морфогенеза («заведующие» формообразованием) функционируют как бы с опережением, задолго до осуществления контролируемых ими формообразовательных событий.
4. Как и предвидел Эдвард Льюис, гомеозисные гены функционируют повсеместно. При этом оказалось, что образуемый их транскриптами «тигровый» рисунок возникает очень рано, до того как четко проявятся признаки самой сегментации. Налицо, таким образом, еще один пример своеобразной химической преформации (предобразования).
6. Гомеобокс и гомеодомен. Принцип коллинеарности
Особо важным открытием было обнаружение в генах, контролирующих сегментацию и превращение, высококонсервативной области ДНК из 180 пар оснований. Эту короткую последовательность Вальтер Геринг назвал гомеобоксом. Соответствующая последовательность из 60 аминокислот в кодируемых этими генами белках была обозначена термином гомеодомен. Гомеодомены являются составной частью белков — регуляторов транскрипции, принадлежат к числу активирующих транскрипцию факторов и характеризуются специфической структурой типа «спираль-поворот-спираль» (рис. 8).
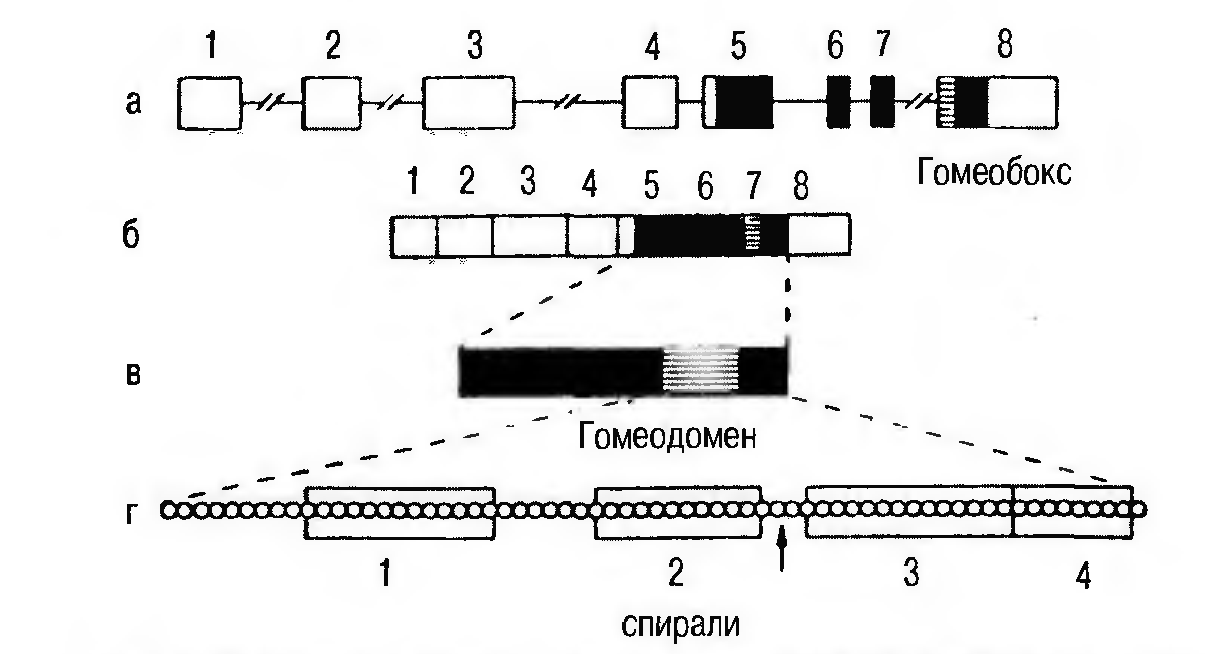
Рисунок 8. Схема строения гена, мРНК и белкового продукта гена Antennapedia: а-ген Antennapedia, б-экзоны (1-8), транскрипты которых образуют мРНК (5-8 – транслируемая область мРНК) в- белок Antennapedia, область гомеодомена, г-структура гомеодомена [1]
Гены, содержащие гомеобокс, были найдены практически у всех живых организмов (губок, пиявок, нематод и др.), они богато представлены и у млекопитающих. Гены, содержащие гомеобокс, которые собраны у представителей этого класса животных в кластер, принято в настоящее время называть НОХ-генами. Они особенно хорошо изучены у мыши и человека. В геноме млекопитающих обнаружено 38 НОХ -генов, собранных в 4 блока. Расположение генов в кластере в основном соответствует расположению гомологичных генов в хромосомах дрозофилы.
Для выяснения функции НОХ-генов в эмбриогенезе млекопитающих использовали метод нокаутирования. С его помощью «выключают» ген, функции которого планируется изучать. Ещё один подход – изучение фнкционирования данного гена в необычном месте. В этом случае изучаемый ген вводят в геном другого животного под промотором (фрагмент ДНК, с которого начинается синтез РНК), который активируется в клетках, где данный ген в норме не функционирует. Необычный промотор заставляет его работать в необычном месте. Что же дало использование этих двух методов для понимания функций гомеозисных генов?
Прежде всего, было выявлено, что у позвоночных, как и у дрозофилы, выполняется принцип коллинеарности, т. е. корреляции между положением генов в кластере и расположением зон их экспрессии (активности) вдоль оси тела. И, во-вторых, было установлено, что гены развития подразделяются на два типа: гены – господа и гены – рабы.