

Палеофлористическое районирование суши
Пространственное распространение определенных совокупностей ископаемых растений позволяет проводить палеофлористическое районирование. При этом используют те же единицы, что и при современном флористическом районировании: царство — область — провинция.
Первые наземные растения силура во всех регионах имели довольно бедный и почти одинаковый состав (риниофиты и плауновидные). Они произрастали на побережьях различных водоемов. Девонские растения намного разнообразнее, чем силурийские. Среди них известны риниофиты, плауновидные, хвощевидные и папоротниковидные, но и девонская флора почти повсеместно имела однотипный состав. Космополитизм девонской флоры хорошо устанавливается по распространению рода Archaeopteris. Эти растения в отложениях верхнего девона встречаются почти повсеместно (Северное полушарие и Австралия), причем именно род Archaeopteris занимает доминирующее положение по отношению к другим растениям (архебптерисовая флора).
С раннего карбона проявляется четкая палеофлористическая дифференциация, позволяющая выделить три области: Гондванскую (Гондвана), Еврамерийскую (Еврамерика) и Ангарскую (Ангарида). В позднем карбоне дополнительно обособилась Катазиатская область, отделение которой от Еврамерийской началось в среднем карбоне (рис. 72). В Еврамерийской и Катазиатской областях в карбоне и перми произрастала тропическая и субтропическая флора экваториального пояса, а в Ангарской (Северное полушарие) и Гондванской (Южное полушарие) областях — флора бореального и нотального умеренно холодных поясов (рис. 73, А). В пределах Еврамерийской области в раннем карбоне (Северная Америка, северная часть Южной Америки, Европа, Кавказ, Средняя и Малая Азия, Северная Африка) произрастали древесные плауновидные (Lepidodendron, Sigillaria, Lepidophloios), хвощевидные (Calamites, Sphenophyllum) и папоротниковидные с крупными вайями. Как правило, преобладали плауновидные (рис. 74). Стволы деревьев имели мощную кору и слаборазвитую древесину (маноксилический тип стебля), что характерно для растительности влажных тропиков и субтропиков (мангровые заросли). Ризофоры лепидодендроновых нередко слагают «стигмариевые почвы». Рост лепидодендроновых на влажных, нередко затопляемых участках отразился на строении стигмарий, у которых гипертрофированное развитие получают воздухоносные полости.
Растительность мангр и болот явилась основой мощного позднепалеозойского углеобразования (Донбасс, Силезия и др.). В среднем карбоне на фоне резкого сокращения видового состава отмечается появление новых групп растений, в том числе и пинофитовых, а с позднего карбона началось сокращение плауновидных (так, в рассматриваемой области перестал произрастать род Lepidodendron). Вместе с тем возросли количество и разнообразие папоротниковидных, в том числе и древовидных. Существенную роль приобретают кордаитовые, о быстром росте которых свидетельствуют находки Artisia. Изменился и климат, в позднем карбоне он стал более сухим, а в перми — полуаридным и аридным. Одновременно с изменением климата в позднем карбоне сократились площади угленакопления. В перми в Еврамерийской области продолжалось сокращение разнообразия плауновидных, уменьшились размеры папоротниковидных. Вместе с этим очень широкое распространение получили пинофитовые.
Катазиатская область (Япония, Корея, Китай, Вьетнам, Лаос, Таиланд, Индонезия, Малайзия), как сказано выше, обособилась от Еврамерийской в позднем карбоне и тоже принадлежала экваториальному поясу. Флора карбона и перми в Катазиатской области близка по составу одновозрастной флоре Еврамерийской области. Произрастали общие роды и виды плауновидных, настоящих папоротников, птеридоспермов, клинолистников и кордаитовых.
Начиная с позднего карбона и еще более резко в перми начинает проявляться специфический облик катазиатской флоры: редки хвойные и кордаитовые. Долго сохраняется преобладание плауновидных, значительно дольше, включая поздний карбон и Пермь произрастает Lepidodendron и вместе с тем полностью отсутствует Sigillaria.
Особенно своеобразный облик катазиатской флоре придавало присутствие, а часто и доминирование в ней уникальных голосеменных — гигантоптерид. Субтропический и тропический климат и меньшая аридизация обусловили накопление угленосных толщ на протяжении ранней перми, т.е. позднее, чем в Еврамерийской области.
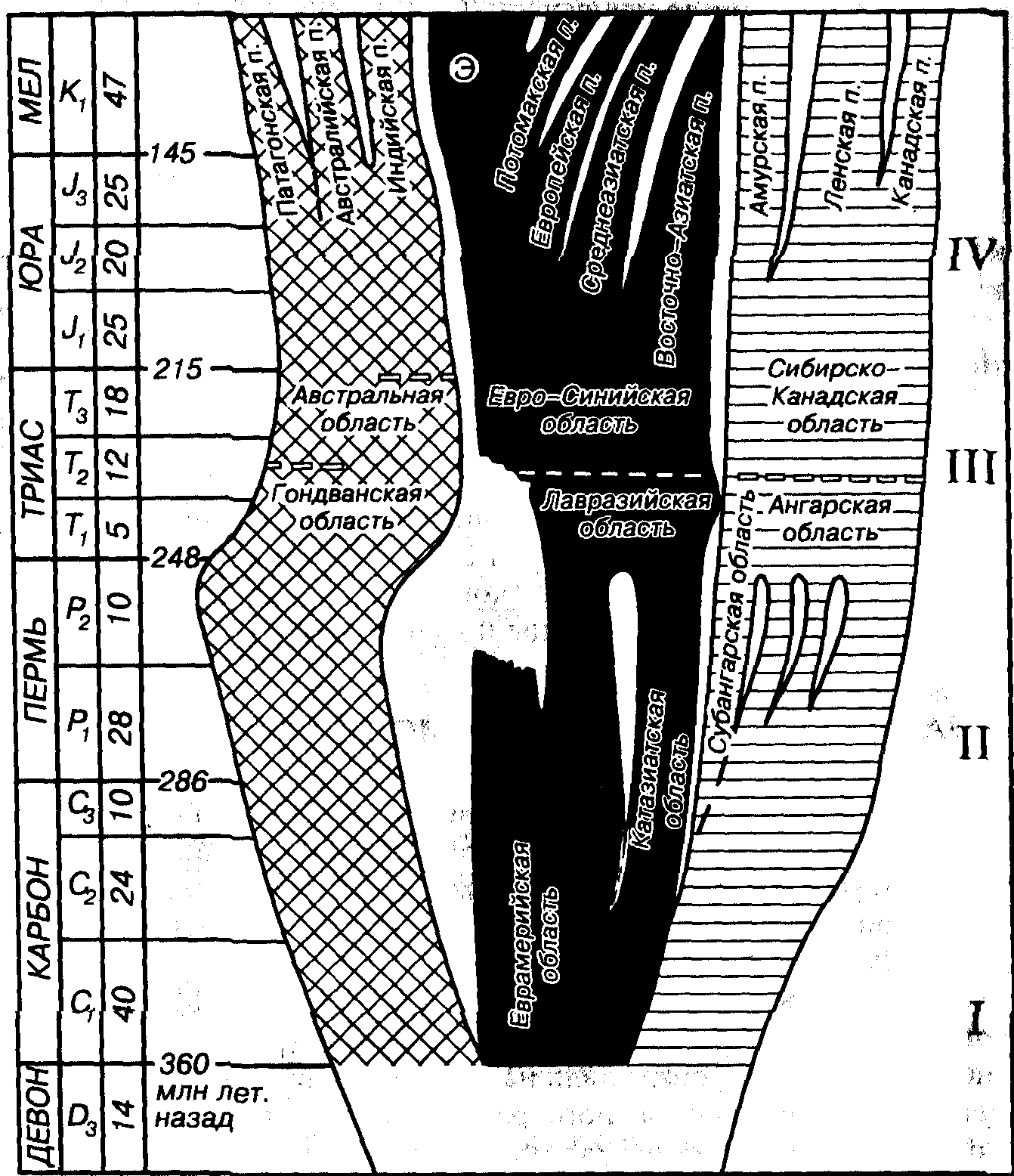
Рис. 72. Эволюция наземных флор в палеозое и мезозое
(Мейен, Вахрамеев, 1984,1987; с упрощением)
Э — экваториальная область; I — дифференциация палеофитной флоры на царства и области; II — последующая дифференциация палеофитной флоры на области и провинции; III — рубежи смены палеофитной флоры на мезофитную флору; IV - последующая дифференциация мезофитной флоры на области и провинции (п.)
Флора Ангарской области (Печора, Сибирь, Казахстан, Монголия) в начале карбона была представлена преимущественно лепидодендроновыми, травянистыми хвощевидными, папоротниковидными со слаборазвитыми вайями, голосеменными. Лепидодендроновые имели небольшие размеры, неветвящийся стебель (рис. 75); примечательно отсутствие стигмарий. В начале карбона климат был безморозным. Об этом свидетельствует маноксилическое строение стебля лепидодендроновых. Такой стебель у многолетних растений мог развиваться только в условиях безморозного климата.
В среднем карбоне началось похолодание, места произрастания раннекаменноугольных плауновидных освоили иные группы растений и в первую очередь кордаитовые (находки артизий отсутствуют). Особенно многочисленными они были с начала перми, составляя основной компонент бореальной «кордаитовой» тайги. В листовой флоре кордаитовых преобладал род Rufloria, поэтому возможно говорить о «руфлориевой тайге». Господство кордаитов подтверждается многочисленными находками листьев, скопления которых образуют «листовые кровли» (термин В.А. Красилова). Одновременно с кордаитовыми произрастали членистостебельные и папоротники. Кордаитовые явились основными углеобразователями (Кузнецкий, Минусинский и Тунгусский бассейны)
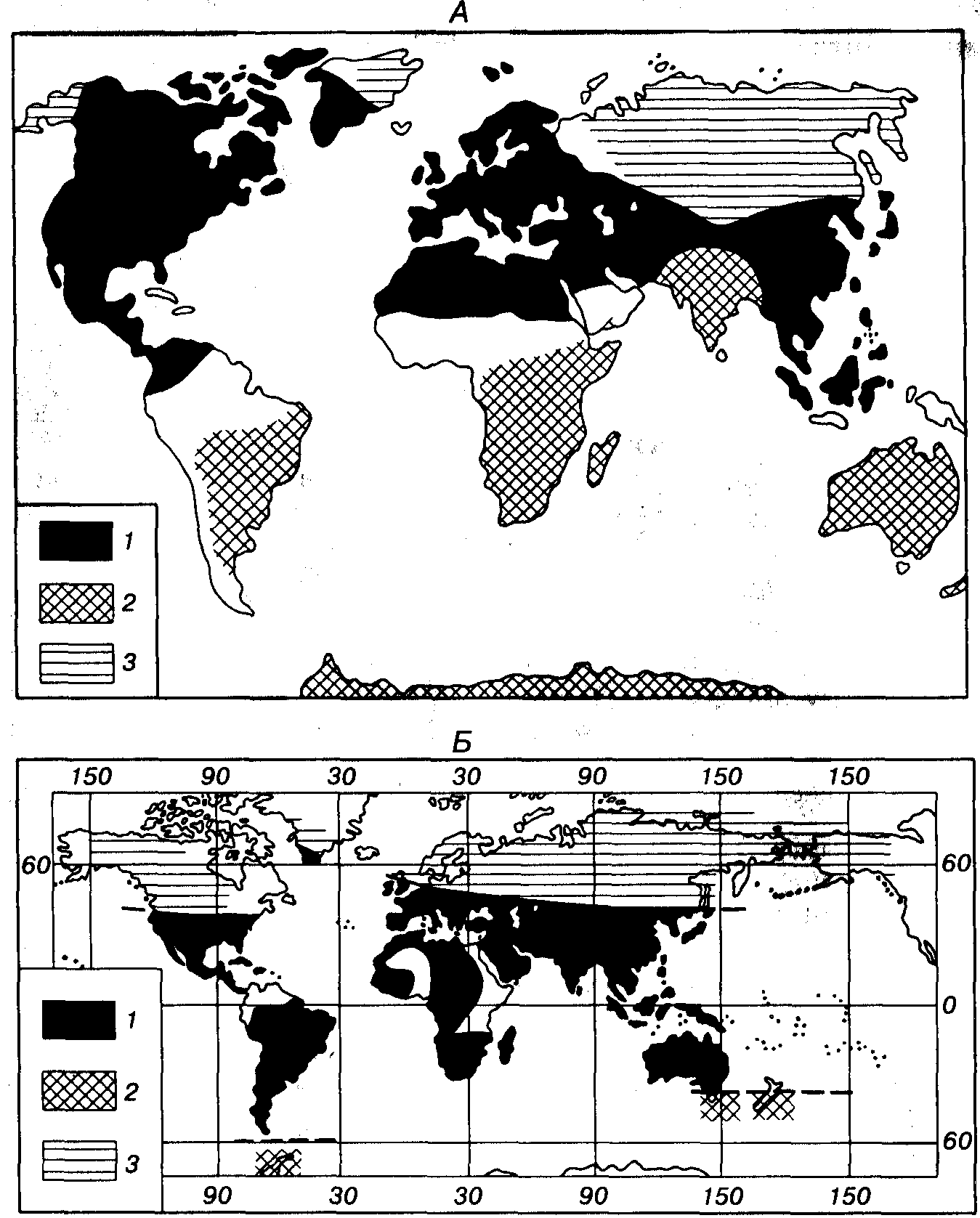
Рис. 73. Фитоклиматическое районирование прошлого А — поздний палеозой (Мейен, 1987); Б — ранний мел (Красилов, 1985, с упро- щением). Флоры: 1 — экваториальные и примыкающие к ним, 2 — нотальные,
3 — бореальные

Рис. 74. Реконструкция среднекаменноугольного ландшафта Северной Англии и
Южной Шотландии (Мейен, 1987)
1 — птеридоспермы (Cycadofilicales), 2 — каламитовые, 3 — настоящие папоротники,
4 — сигиллярии, 5 — лепидодендроны

Рис. 75. Реконструкция раннекаменноугольного ландшафта Сибири (Мейен, 1988)
Гондванская область, занимавшая огромные пространства (Южная Америка, Южная Африка, Индия, Австралия и Антарктида), представляла собой единый материк — Гондвану, площадь которого занимала не менее трети современной суши. Эта область располагалась в нотальном поясе и характеризовалась умеренным или умеренно прохладным климатом. Видимо, климат в целом был более холодным, чем в Ангарской области. Флора была представлена тонкоствольными плауновидными, папоротниками, близкими к листовой флоре Еврамерийской области, но существенно отличающимися по споровым комплексам. Принципиальные отличия гондванской и еврамерийской флор проявляются начиная с позднего карбона.
С этого времени фиксируются появление и массовое развитие глоссоптерид. Господство глоссоптерид, известных повсеместно, подтверждается скоплениями листьев («листовые кровли»), подобно тому, что наблюдалось в ангарской флоре для листьев кордаитовых. Глоссоптериевая флора характеризуется преобладанием родов Gangamopteris и Glossopteris.
К концу карбона исчезли крупные древовидные плауно- и хвощевидные. Как в Ангарской области, отсутствуют стигмарии. С позднего карбона на территории Гондваны началось оледенение, продолжавшееся и в ранней перми. Ледник оставил мощные толщи тиллитов — ледниковых глинистых отложений с валунами. Периоды похолодания сменялись периодами потепления. Климат был влажным, формировались мощные пласты углей. В конце перми началась аридизация климата, продолжавшаяся в раннем триасе.
Перестройка растительного мира началась в конце перми. Многие группы растений вымерли, на смену им пришли новые растения и возникли новые ассоциации, возросла дифференциация растительности. В триасе границы между Гондванской, Лавразийской (Еврамерийская плюс Катазиатская) и Ангарской областями становятся менее отчетливыми. В среднем триасе завершается глобальное обновление флор и палеофитная флора сменяется мезофитной. С этого момента выделяются области: Евро-Синийская, сменившая Еврамерийскую, Сибирско-Канадская, трансформировавшаяся из Ангарской, и Австральная (Австралийская), преобразованная из Гондванской. В различных областях отдельные элементы и ассоциации мезофитной флоры появлялись в разное время.
В Евро-Синийской области был влажный теплый климат экваториального пояса. Появилось много древовидных папоротниковидных, голосеменных (цикадовых и беннеттитовых), образующих леса тропиков и субтропиков. Хвойные играли резко подчиненную роль. Максимальный расцвет этой флоры приурочен к юре, что привело к мощному углеобразованию. В Евро-Синийской области в начале мела возникли первые цветковые растения.
В Сибирско-Канадской области климат был умеренно холодный. Тайгу в основном образовывали гинкговые, чекановскиевые, хвойные (древние сосновые). Названные растения имели пикноксилический стебель, для которого характерно резкое преобладание древесины над корой. Цикадовые и беннеттитовые здесь отсутствовали. В это же время в Австральной, или Австралийкой, области наблюдается много цикадовых и беннеттитовых, достигавших в высоту 15 м и более, немногочисленны хвойные имеющие не игольчатые, а ланцетовидные листья.
В юре, позднем мелу и палеогене дифференциация растительности еще более усилилась; возникли 11 палеофлористических провинций, но границы между областями стали менее четкими. Начиная с неогена формируются современные фитогеографические царства, области, провинции, округа. Кайнофитная флора сменила мезофитную.