

Отдел плауновидные. Divisio lycopodiophyta
(греч. lykos — волк; pous, podos — нога; phyton — растение)
Плауновидные — наземные растения, имеющие стебель, филлоидные листья и корни (или ризофоры). В современной флоре известны только травянистые формы, к которым относится собственно род Lycopodium — плаун. Гаметофит современного плауна существует в виде самостоятельного подземного растения, от которого начинается рост спорофита. На рис. 42 показано, что споры плауна, попадая в почву (прорастают на 3-8-й год), дают начало гаметофиту (существует до 12 лет, а затем отмирает). От погруженного в землю гаметофита возникает зародыш споро- фита. Естественно, что этому предшествует возникновение в верхней части гаметофита архегониев и
зрелый спорофит

антеридиев с половыми продуктами. Цикл развития спорофит — гаметофит — новый спорофит подобен таковому у папоротников (см. рис. 38).
Первые плауновидные, произошедшие в позднем силуре от риниофитов, были небольшими травянистыми растениями. В позднем палеозое появляются крупные древовидные формы. Возникновение деревьев как таковых стало возможным благодаря появлению камбия, что определило вторичный рост растений. В стволе, как правило, резко преобладает кора, составляющая у древесных форм до 80-90% (порядок Lepidodendrales).
Нижняя подземная часть ствола обычно многократно дихотомически ветвилась, образуя так называемый ризофор (= стигмарии). Ризофор (греч. rhiza — корень; phoros — несущий) по положению соответствует корням, хотя, возможно, он мог быть погружен в субстрат неполностью. От ризофора отходили аппендиксы, при их отпадании оставался небольшой округлый отпечаток «глазок», или стигма, в центре которого наблюдается выход проводящего пучка. По анатомическому строению ризофор и аппендиксы отличаются от настоящих корней и корневых волосков.
Примитивность плауновидных проявляется в строении листоподобных образований филлоидов. У наиболее примитивных форм имелись только эмергенцы. Филлоиды обычно узкие, удлиненные, с листовым следом, но еще не имевшие листовых прорывов (см. рис. 37). Иногда листья плауновидных по морфологии с трудом отличимы от подобных образований хвойных (сходство конвергентное). Проводящая система построена по варианту актинос- телы (разновидность протостелы) либо сифоностелы. Споры равных или разных размеров, они образуются в спорангиях, которые располагаются на верхней стороне или в пазухах видоизмененных листьев — спорофиллов. Последние нередко группируются в стробилы (греч. strobilos — сосновая или еловая шишка). В одном стробиле могли находиться и макро- и микроспорофиллы (см. рис. 44).
При разделении плауновидных на порядки в первую очередь используются два признака: 1) равно- или разноспоровость, 2) наличие или отсутствие лигулы (лат. ligula — язычок), представляющее собой особое образование при основании листа или в его пазухе. Лигула иногда помещается в углублении (см. рис. 46). Ее функция недостаточно ясна. Равноспоровые плауновидные, за редким исключением, не имели лигулу, а у разноспоровых она была. Ниже рассмотрены только три порядка: Drepanophycales (S2?, D), Protolepidodendrales (D1_2) и Lepidodendrales (C-T).
Порядок Drepanophycales охватывает наиболее древних плауновидных, которые, видимо, возникли от риниофитов в позднем силуре. Это были кустарниковые растения, не превышающие в высоту 1 м (рис. 43). Стела устроена просто: актиностела, имеющая звездчатую форму на поперечном сечении, в нижней части ствола иногда образуется сифоностела. Стебель дихотомически ветвящийся. Мелкие, на концах игольчатые, филлоиды или эмергенцы обычно располагались по спирали. Лигула отсутствовала. У рода Drepanophycus проводящий пучок заходил в филлоидный лист, а у рода Asteroxylon проводящий пучок доходил только до основания эмергенца, не заходя внутрь (см. рис. 43, в). Округлые или удлиненные спорангии находились на верхней поверхности спорофиллов либо были углублены в их пазухах или прикреплялись к стеблю. Споры равных размеров. Надземная стелющаяся часть стебля ориентирована горизонтально.
Дрепанофиковые известны из отложений девона, видимо, они появились в позднем силуре. Иногда с ними сближают кембрийский род Aldanophyton. Находки этого рода приурочены к битуминозным известнякам, которые могли образовываться только вдали от берега, и поэтому сомнительно, «сто Aldanophyton мог

Рис. 43. Порядок Drepanophycales (S2?, D)
а-г — Asteroxylon (D2): a — реконструкция, б — поперечный разрез стебля, в - продольный разрез стебля с выступами — эмергенцами, г — реконструкция побе- га со спорангиями; д, е — Drepanophycus (D): д — реконструкция, е — продоль- ный разрез стебля с филлоидами
 принадлежать
высшим растениям. Сейчас начинают
оспаривать и силурийские находки
дрепанофиковых, тем самым ограничивая
интервал их существования только
девоном.
принадлежать
высшим растениям. Сейчас начинают
оспаривать и силурийские находки
дрепанофиковых, тем самым ограничивая
интервал их существования только
девоном.
Порядок Protolepidodendrales (греч. protos — первый; Lepidodendrales — название одного из порядков плауновидных) включает небольшую группу ранне-среднедевонских травянистых растений высотой до 20-30 см (рис. 44). Характерно наличие подземного горизонтального корневища, от которого отходили вертикальные дихотомически ветвящиеся побеги. Вильчатые или пятилопастные филлоиды имеют преимущественно спиральное расположение. При отпадении филлоидов оставались возвышения — зачаточные листовые подушки. У большинства форм была лигула — язычок. Спорангии располагались на верхней поверхности филлоидов, спорофиллы могли группироваться вместе, занимая на побеге конечное положение. Споры равных размеров. Протолепидодендроновые дали начало следующему порядку, с которым их сближают однотипное расположение листьев и наличие зача- точных листовых подушек.
Порядок Lepidodendrales (греч. lepis, lepidos — чешуя; dendron — дерево) представлен крупными вымершими древесными растениями, которые достигали в поперечнике 2 м и в высоту 30-40 м (рис. 45). Стволы лепидодендроновых нередко дихотомически ветвились, создавая широкую крону. Удлиненные, обычно линейные

Рис. 44. Отдел Lycopodiophyta (S2-Q)
а--в — Protolepidodendron (Di_2): a — реконструкция, 6 — часть стебля с филлоидами, в — филлоид со спорангиями; г — схема строения стробила рода Flemingites (C-P)
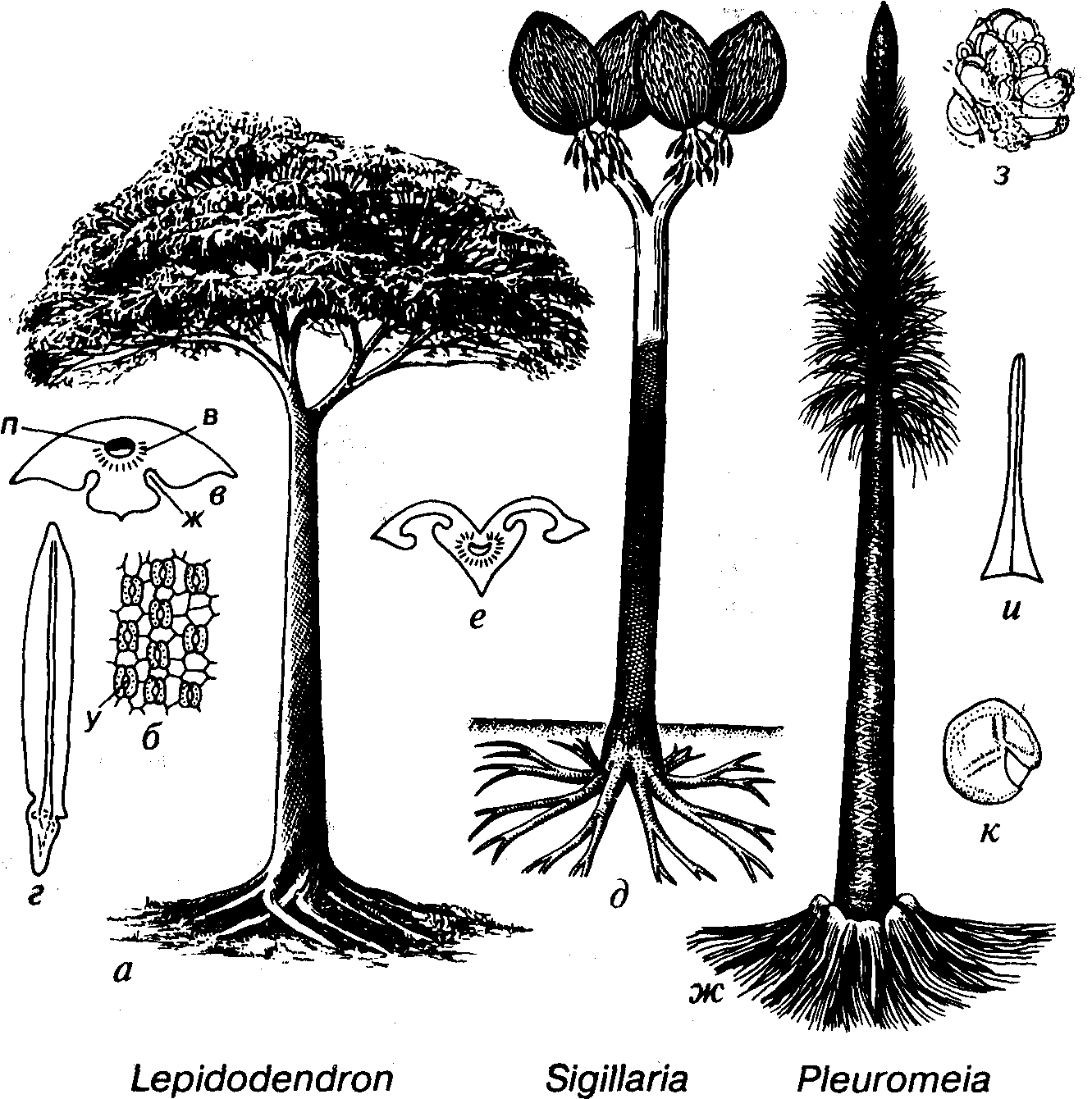
Рис. 45. Порядок Lepidodendrales (C-T)
а-в — Lepidodendron (C-P): а — реконструкция, б — устьица в желобках филлоида, в — поперечное сечение филлоида; г — Lepidostrobophyllum (C2), спорофилл; д, е — Sigillaria (C-P): д — реконструкция, е — поперечное сечение филлоида; ж-к — Pleuromeia (T): ж — реконструкция, з — стробил, и — филлоид, к — микроспора, в — воздухоносная ткань, ж — желобок с устьицами, п — проводящий пучок, у — устьица
филлоиды, иногда имеющие длину до 1 м, закономерно располагались на приподнятых участках ствола, получивших название листовых подушек. По мере роста дерева и опадания листьев ствол обнажался и на коре проступал правильный рельеф листовых подушек. Листовая подушка рода Lepidodendron имеет форму ромба, вытянутого в высоту (рис. 46). В ее верхней трети на месте отпадания, листа обособился листовой рубец. Листовой рубец по форме был почти квадратными либо подобно листовой подушке имел форму ромба, но вытянутого по горизонтали. В пределах листового рубца выделяются три элемента. Средний, листовой, след является отпечатком
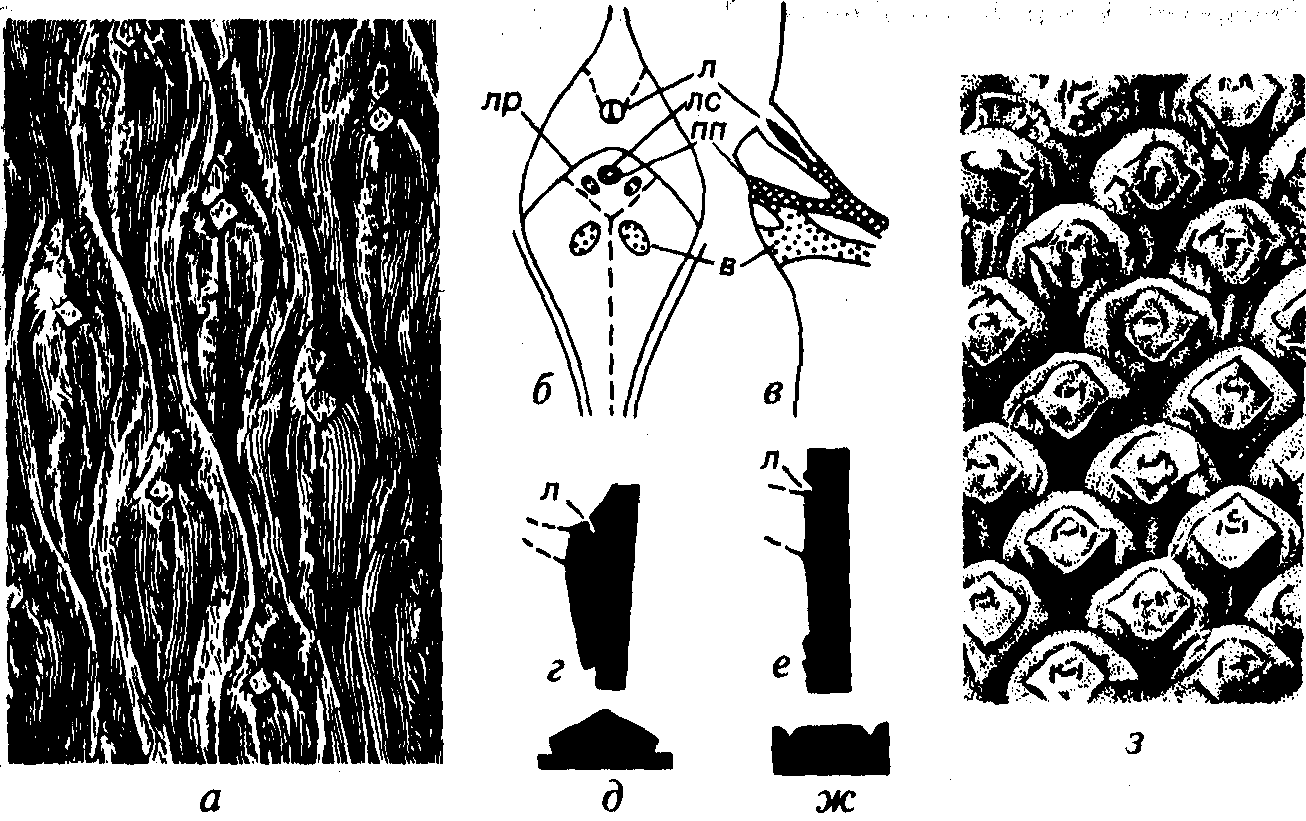
Рис. 46. Порядок Lepidodendrales (C-T) а-д — Lepidodendron (C-P): а — отпечаток коры с листовыми подушками, б, в — схема строения листовой подушки (б — анфас, в — продольный разрез), г, д — контур листовой подушки вдоль и поперек стебля; е-з — Sigillaria (C-P): е-ж —контур листовой подушки вдоль и поперек стебля, з — кора с листовыми подушками, в —воздухоносная ткань, л — лигула, лр — листовой рубец, лс — листовой след, пп — проводящий пучок (г, д, е, ж — Мейен, 1987)
проводящего пучка, а боковые — представляют выходы двух воздухоносных тяжей. Два подобных тяжа располагались ниже листового рубца за его пределами. Воздухоносные тяжи способствовали газообмену. В верхней части листовой подушки располагалась лигула. Листовые подушки лепидодендроновых по мере роста растения разрастались и увеличивались, поэтому в нижней части дерева они в 20-30 раз превосходят таковые в верхней его части, где листья только отпали.
У некоторых лепидодендроновых, например у рода Sigillaria (см. рис. 46), листовые подушки отсутствовали и листовые рубцы, подобные таковым у рода Lepidodendron, могли располагать- ся на ребристой или гладкой коре.
Степень сохранности коры различна, поэтому форма и строение листовых подушек будут существенно различаться, что схематически показано на рис. 47. Для различных форм сохранности предложены самостоятельные латинские названия (роды Bergeria, Knorria, Helenia).
Ризофор обычно состоит из четырех главных ветвей, которые многократно дихотомически разделялись и неглубоко погружались в землю (рис. 48). У высоких древесных форм площадь, занятая ризофором, могла в диаметре превышать 20 м. У триасового рода Pleuromeia подземная часть растения имеет форму расширенного клубня с четырьмя якореподобными отростками.
Лепидодендроновые были разноспоровыми растениями, для мегаспор и микроспор характерна трехлучевая щель разверзания (см. рис. 45, к). Спорофиллы собраны в стробилы, которые могли содержать как микро-, так и мегаспорангии (см. рис. 44). У некоторых плевромей отсутствовали стробилы, а имелись только фертильные зоны со спорангиями.
Ствол лепидодендроновых имел толстую кору и тонкий проводящий пучок, устроенный преимущественно по типу прото- или сифоностелы. Такое соотношение коры и проводящего пучка характерно для стволов маноксилического типа (греч. manos — редкий, неплотный; xylon — древесина, срубленное дерево). Такие стволы можно называть малодревесинными. Растения с

Рис. 47. Схема строения коры и стебля лепидодендроновых
а — блок-диаграмма, показывающая различные формы сохранности коры и древесины, б — схема расположения тканей на поперечном срезе стебля, дп, дт — ксилема (древесина) первичная и вторичная; кв, кн — первичная кора внутренняя и наружная; ло, лт — лучи основной и механической тканей; пв, пн — перидерма внутренняя и наружная; ел — листовой след; слп — листовые подушки
маноксилическим стеблем произрастали в условиях безморозного климата. В тех случаях, когда в стволе лепидодендроновых помимо толстой коры присутствовали хорошо развитые воздухоносные ткани, можно предполагать, что такие формы росли в тропических и субтропических условиях. Возможно, что нижняя часть растений периодически затоплялась.

Рис. 48. Порядок Lepidodendrales (C-T) а, б — Stigmaria (C-P): а — дихотомически ветвящийся ризофор, б — фрагмент ризофора с аппендиксами; в — Protostigmaria (С1), клубневидный ризофор с ап- пендиксами; г — Pleuromeia (T), нижняя часть ствола с клубневидным ризофором, видны рубцы листьев-филлоидов и аппендиксов
В ископаемом состоянии от лепидодендроновых сохраняются отпечатки коры, листьев, стволы, изредка органы спороношения. Как правило, находки бывают разрозненны и получают самостоятельные латинские названия, а предлагаемые реконструкции нередко спорны, ибо далеко не всегда представляется возможность доказать, что все составные части в действительности принадлежали одному растению. Так, Lepidostrobus — шишка лепидодендроновых, a Stigmaria — подземная часть стебля.
Лепидодендроновые, вероятно, возникли в позднем девоне, их расцвет приурочен к карбону, в триасе они вымерли. Они составляют значительную часть углей карбона и перми.