

Нормальная физиология (Пособие для резидентуры)-1
.pdfОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ |
2 |
Сравнительнаяхарактеристикаместногоираспространяющегося возбуждения
Местное возбуждение /4/:
 возникает при действии слабых подпороговых стимулов;
возникает при действии слабых подпороговых стимулов;
 возникает локально – только в зоне действия стимула;
возникает локально – только в зоне действия стимула;
 быстро затухает во времени (распространяется с декрементом);
быстро затухает во времени (распространяется с декрементом);
 не распространяется на значительное расстояние;
не распространяется на значительное расстояние;
 способно к суммации локальных токов и может перейти в распространяющееся возбуждение;
способно к суммации локальных токов и может перейти в распространяющееся возбуждение;
 графически характеризуется незначительным отклонением уровня потенциала покоя (предспайк);
графически характеризуется незначительным отклонением уровня потенциала покоя (предспайк);
 характеризуется некоторым повышением возбудимости ткани;
характеризуется некоторым повышением возбудимости ткани;
 не подчиняется закону «все или ничего».
не подчиняется закону «все или ничего».
Распространяющееся возбуждение:
 возникает при действии пороговых и надпороговых стимулов, вследствие суммации локальных токов;
возникает при действии пороговых и надпороговых стимулов, вследствие суммации локальных токов;
 распространяется на значительное расстояние;
распространяется на значительное расстояние;
 не «затухает» во времени (бездекрементно);
не «затухает» во времени (бездекрементно);
 не способно к суммации;
не способно к суммации;
 графически характеризуется развитием потенциала действия (спайка) и инверсией заряда мембраны;
графически характеризуется развитием потенциала действия (спайка) и инверсией заряда мембраны;
 подчиняется закону «все или ничего»;
подчиняется закону «все или ничего»;
 сопровождается циклическими изменениями возбудимости и наличием периода абсолютной рефрактерности;
сопровождается циклическими изменениями возбудимости и наличием периода абсолютной рефрактерности;  имеет большое физиологическое значение, особенно при реализации «быстрых»
имеет большое физиологическое значение, особенно при реализации «быстрых»
процессов, например, проведении возбуждения в двигательных соматических нервах.
Следовыепотенциалы
Следовые потенциалы впервые были зарегистрированы Д.С. Воронцовым, а дальнейшем подробно изучены Д. Эрлангером и Г.С. Гассером.
Различают два вида следовых потенциалов – следовую деполяризацию (следовой отрицательный потенциал) и следовую гиперполяризацию (следовой положительный потенциал) (рис. 2.8). Амплитуда этих потенциалов не превышает нескольких милливольт, а длительность варьирует от нескольких миллисекунд до несколько десятков или даже сотен миллисекунд. Следовые потенциалы связаны с восстановительными процессами, медленно развивающимися в нервных и мышечных волокнах после окончания возбуждения.
Взаимоотношения между пиком потенциала действия и фазой следовой деполяризации могут быть рассмотрены на примере ответа скелетного мышечного волокна (рис. 2.12), где видно (рис. 2.12), что нисходящая фаза потенциала действия делится на две неравные части. Вначале падение потенциала действия происходит быстро (б), а затем сильно замедляется. Этому моменту и соответствует начало следовой деполяризация (в).
31

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
мВ |
|
+30 |
|
0 |
|
а |
б |
–50 |
|
|
в |
–100 |
|
мсек |
|
Рис. 2.12. Потенциал действия скелетного мышечного волокна, зарегистрированный с помощью внутриклеточного микроэлектрода: а – фаза деполяризации; б – фаза реполяризации; в – следовой отрицательный потенциал. Момент нанесения раздражения показан стрелкой /1/.
Следовая гиперполяризация мембраны выражается в увеличении исходного потенциала покоя, которая хорошо выражена в безмякотных нервных волокнах. Так, в безмякотном гигантском аксоне кальмара нисходящая фаза потенциала действия непосредственно переходит в фазу следовой гиперполяризации, амплитуда которой достигает примерно 15 мв (рис. 2.13) /1/. В миелинизированных нервных волокнах следовые изменения потенциала имеют более сложный характер: следовая деполяризация сменяется следовой гиперполяризацией, затем иногда развивается новая деполяризация, и лишь после этого происходит полное восстановление потенциала покоя. При ритмическом раздражении нерва следовые потенциалы суммируются вследствие чего их амплитуда и длительность возрастают.
мВ
+40
0
–70 |
|
Q |
|
а
мс
б
Рис. 2.13. Потенциал действия гигантского аксона кальмара, записанный с помощью внутриклеточного микроэлектрода (по Ходжкину и Хаксли). Выражен следовой положительный потенциал (а). отметка времени (б) – 500 колебаний в 1 секунду.
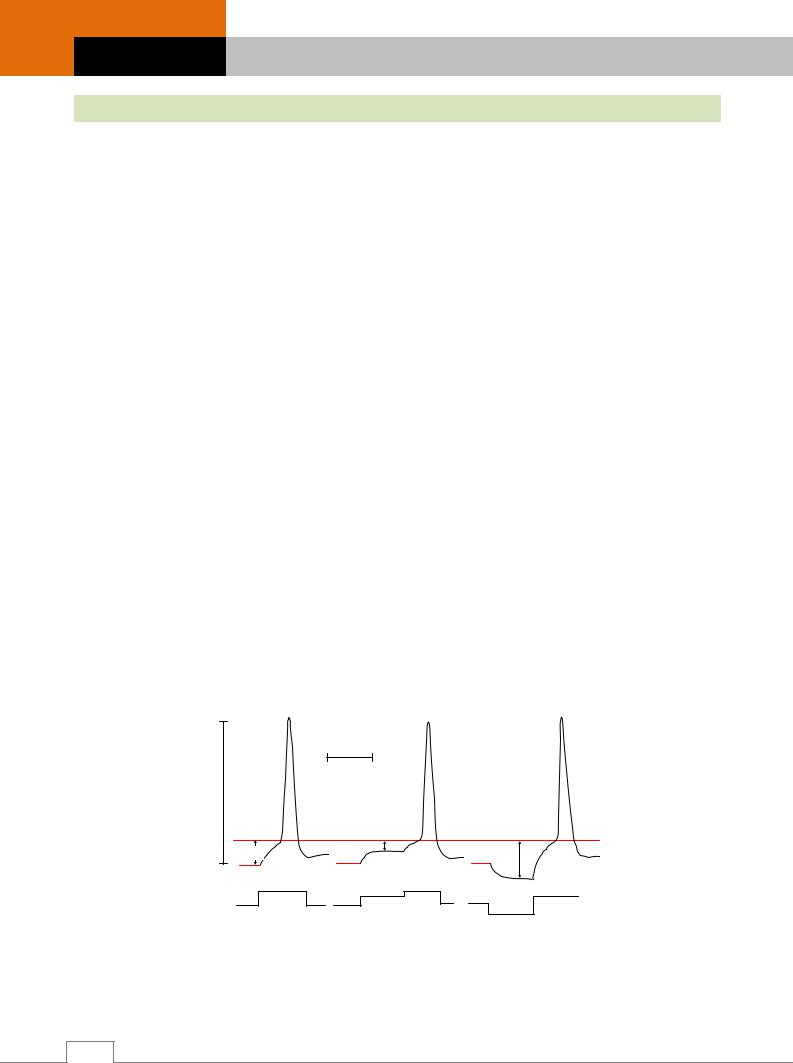
Изменениевозбудимостивразличныефазыодиночного циклавозбуждения
В динамике развертывания одиночного цикла возбуждения происходят изменения возбудимости ткани (рис. 2.14) /4/.
Если принять уровень возбудимости в условиях физиологического покоя за норму, то в ходе развития одиночного цикла возбуждения можно наблюдать ее циклические колебания.
Во время предспайка разница между уровнем заряда мембраны и уровнем критической деполяризации уменьшается. Появляется возможность дополнительным пороговым раздражением меньшей величины быстрее открыть поры для ионов. Na+, достичь уровня критической деполяризации мембраны и быстрее вызвать ПД. Возбудимость, как обратная величина порога, при этом увеличивается и становится супернормальной /4/.
32

ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ |
2 |
V, мВ |
С |
|
|
||
0 |
t, мс |
|
ОСП |
||
|
||
ПП |
|
|
ПС |
ПСП |
|
А |
||
Возбудимость, % |
Э |
|
100 |
||
|
||
СуперВ |
СубВ |
|
|
ОР |
0
БАР
Рис. 2.14. Изменение возбудимости при одиночном возбуждении: А – потенциал действия; Б – динамика возбудимости
(V – напряжение на внутренней поверхности мембраны; t – время; ПП – потенциал покоя;
КД – критическая деполяризация;
Р – раздражение; ПС – предспайк; С – спайк; ОСП – отрицательный следовой потенциал;
ПСП – положительный следовой потенциал;
В – возбудимость; СуперВ – супернормальная возбудимость; АР – абсолютная рефрактерность; ОР – относительная рефрактерность;
Э – экзальтация; СубВ – субнормальная возбудимость).
Во время спайка открываются все поры для ионов Na+ и натрий поступает внутрь клетки. Поэтому никакая величина раздражения не способна усилить процесс возбуждения. Порог раздражения увеличивается до бесконечности, а возбудимость, как обратная величина, падает до нуля – наступает период абсолютной рефрактерности или полной невозбудимости /4/. Свойство возбуждаться исчезает, так как мембрана уже возбуждена. Фармакологические вещества, увеличивающие время деполяризации и соответственно период абсолютной рефрактерности, вызывают торможение физиологической деятельности. Наоборот, вещества и воздействия, уменьшающие время деполяризации, оказывают возбуждающее физиологическое действие. Застойная деполяризация наблюдается, например, при парабиозе. Самый короткий период абсолютной рефрактерности характерен для нерва.
Во время реполяризации поры для ионов Na+ постепенно закрываются, поэтому появляется возможность их заново открыть дополнительным раздражением /4/. При этом сила раздражения должна быть выше величины исходного порогового раздражения. Время восстановления возбудимости от нуля до исходной величины называется периодом относительной рефрактерности. В это время ткань может ответить возбуждением только на сильные, надпороговые раздражения.
Во время отрицательного следового потенциала разница между уровнем заряда мембраны и уровнем критической деполяризации меньше, чем в состоянии покоя /4/. Тогда порог раздражения меньше, а возбудимость больше – стадия экзальтации.
Во время положительного следового потенциала разница между уровнем заряда мембраны и уровнем критической деполяризации увеличивается, порог раздражения увеличивается, а возбудимость уменьшается – наступает период субнормальной возбудимости, совпадающий с развитием гиперполяризации мембраны. Фармакологические вещества и воздействия, усиливающие ее, вызывают ослабление физиологической деятельности (гиперполяризационное торможение). Возбуждение может возникнуть только в том случае, когда сила раздражения значительно выше пороговой.
После завершения указанных процессов возбудимость восстанавливается и клетка готова к осуществлению следующего цикла.
33
НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
2.2. Особенностидействияпостоянноготоканаткани
Закон полярного действия постоянного тока (Э.Пфлюгер): при замыкании тока воз-
буждение возникает под катодом, при размыкании под анодом. Полярный закон доказывается следующими опытами. Умерщвляют участок нерва под одним из электродов, а второй электрод устанавливают на неповрежденном участке. Если с неповрежденным участком соприкасается катод, возбуждение возникает в момент замыкания тока; если же катод устанавливают на поврежденном участке, а анод – на неповрежденном, возбуждение возникает только при размыкании тока. Порог раздражения при размыкании, когда возбуждение возникает под анодом, значительно выше, чем при замыкании, когда возбуждение возникает под катодом. Это связано с тем, что прохождение постоянного тока через возбудимую ткань вызывает изменение мембранного потенциала покоя ее клеток. Так, в области приложения к возбудимой ткани катода положительный потенциал на наружной поверхности клеточной мембраны уменьшается и возникает начальная деполяризация мембраны, которая под влиянием проходящего электрического тока быстро достигает критического уровня и вызывает распространяющееся возбуждение. В области приложения анода положительный потенциал на наружной поверхности мембраны возрастает, происходит гиперполяризация мембраны. При размыкании цепи тока гиперполяризация мембраны исчезает, потенциал возвращается к исходной величине и достигает смещенного критического уровня деполяризации – возникает возбуждение.
Изучение механизма полярного действия постоянного тока стало возможным благодаря методу одновременного введения в клетку двух микроэлектродов: одного – для раздражения, другого – для отведения потенциалов. Было установлено, что потенциал действия возникает только в том случае, когда катод находится снаружи, а анод – внутри клетки. При обратном расположении полюсов, т.е. наружном аноде и внутреннем катоде, возбуждение при замыкании тока не возникает, как бы силен он ни был.
Закон физиологического электротона (Э.Пфлюгер): действие постоянного тока на ткань сопровождается изменениями ее возбудимости. Начальное прохождение постоянного тока через нерв или мышцу в участке ткани, расположенном под катодом повышает возбудимость (катэлектротон), а под анодом – понижает (анэлектротон). Изменения возбудимости обуславливаются частичной деполяризацией клеточной мембраны под катодом и гиперполяризацией под анодом. Эти изменения возбудимости получили название электро-
тона (рис. 2.15) /1/.
А |
Б |
|
В |
|
5 мс |
|
|
100 мВ |
|
|
|
Ек |
Ек |
Ек |
|
Е0 ∆V0 |
∆V1 |
|
|
Е0 |
Е0 |
∆V2 |
Рис. 2.15. Соотношение между исходным уровнем потенциала покоя (Е0), критическим уровнем деполяризации (Ек) и пороговым потенциалом (∆V), в норме (А) и при электротоне (Б, В).
Б – катэлектротоническое снижение порога ∆V1 наступает в результате приближения потенциала покоя Е0 к критическому уровню деполяризации Ек; В – анэлектротоническое повышение порога ∆V2 является следствием удаления исходного уровня потенциала покоя Е0 от Ек.
Нижняя линия – раздражающий и поляризующий токи: катодный ток – вверх, анодный ток – вниз.
34

ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ |
2 |
При длительной деполяризации мембраны развиваются процессы, повышающие критический уровень деполяризации. Такими процессами являются инактивация натриевых каналов и активация калиевых. Рост Ек при данном значении Е0 ведет к увеличению порогового потенциала, т.е. снижению возбудимости (рис. 2.16) /1/. Одновременно с увеличением порога происходит снижение потенциала действия – крутизна его нарастания и амплитуда падают. Такое снижение возбудимости нервного волокна при длительной и сильной деполяризации мембраны было описано Б.Ф.Вериго и получило название катодической депрессии.
мВ |
|
|
|
|
|
|
|
|
|
|
|
Рис. 2.16. Катодическая депрессия Вериго: |
100 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 – ответ нормального нервного волокна на |
|
|
|
|
|
|
|
|
|
|
|
|
пороговый раздражающий ток; 2 – ответ того же |
|
|
|
1 |
|
|
|
|
|
|
волокна через 10 с после начала действия катода |
||
50 |
|
|
|
|
|
|
|
|
подпорогового постоянного тока; Е0, Ек и |
|||
|
|
|
|
|
|
|
Ек1 |
2 |
||||
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
∆V – исходные величины потенциала покоя, |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
критического уровня деполяризации и порогового |
|
|
Ек |
|
|
|
|
|
Е01 |
∆V1 |
|
|
||
|
|
|
|
|
|
|
|
|
|
|
потенциала. Е10, Е1к и ∆V1 – величины тех же |
|
|
Е0 |
∆V |
|
|
||||||||
|
|
|
|
|
|
|
|
|||||
0 |
|
|
|
|
|
|
|
параметров во время катодической депрессии. |
||||
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
1 мc |
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|||||
Описанные явления демонстрируют наблюдающиеся в естественных условиях изменения возбудимости нервных клеток и пресинаптических нервных терминалей при действии на них медиаторов, вызывающих кратковременную или длительную деполяризацию мембраны. В частности, явление катодической депрессии лежит в основе пресинаптического торможения, наблюдающегося в ЦНС.
При длительной гиперполяризации мембраны (в нервном волокне она появляется в области приложения анода) происходит снижение калиевой проницаемости и ослабление исходной натриевой инактивации. Эти изменения ведут к снижению V, возросшего в начальный момент гиперполяризации мембраны, и увеличению амплитуды и крутизны нарастания потенциала действия. Если гиперполяризующий ток прикладывается к мембране, которая предварительно была подвергнута воздействию избытка ионов К+ или действию анестетиков, усиливающих исходную натриевую инактивацию (т.е. увеличивающих долю каналов, у которых инактивационные «ворота» закрыты), то во время длительной гиперполяризации V может снизиться настолько, что при выключении тока, т.е. при возвращении мембранного потенциала к его исходной величине, возникает потенциал действия (рис. 2.17).
б
а
Ек Ек
Е0 Е0 
Рис. 2.17. Возникновение анодно-размыкательного возбуждения: а – потенциал действия, возникающий под катодом при замыкании тока; б – потенциал действия, возникший под анодом при размыкании сильного тока. Стрелкой вниз показано включение анодного тока, стрелкой вверх – его выключение /1/.
35

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
Такое явление получило название анодно-размыкательного возбуждения. В некоторых нервных клетках на подобном механизме основано возникновение потенциала действия после окончания тормозного гиперполяризационного потенциала в естественных условиях. По-видимому, у таких клеток инактивация натриевых каналов и активация калиевых сильно выражены при потенциале покоя в нормальной солевой среде.
Изложенные данные о влиянии деполяризации и гиперполяризации мембраны на ее возбудимость представлены в табл. 2.1 /1/.
Таблица2.1. Измененияфизиологическихпараметровмембраныпридеигиперполяризации
Физиологические |
Деполяризациямембраны |
Гиперполяризациямембраны |
параметры |
(катэлектротон) |
(анэлектротон) |
|
|
|
Возбудимость |
Вначале повышение, затем понижение |
Сначала понижение, затем |
|
|
относительное повышение |
|
|
|
Потенциал действия |
Понижение, прогрессирующее по |
Повышение, прогрессирующее |
|
времени вплоть до полного угнетения |
во времени |
|
|
|
Скорость проведения |
Вначале повышение, затем понижение |
Вначале понижение вплоть до |
|
вплоть до полного блока |
блока (при сильном токе), затем |
|
|
постепенное восстановление |
|
|
|
Натриевая |
Вначале повышение, приводящее к |
Постепенное ослабление инакти- |
проницаемость |
возникновению при подпороговом токе |
вации, если она имела место |
|
локального ответа, а при пороговом – |
|
|
потенциала действия, затем |
|
|
постепенная инактивация |
|
|
|
|
Калиевая проницаемость |
Постепенное повышение |
Понижение, если она была |
|
|
повышена |
|
|
|
В лабораторных условиях и при проведении некоторых клинических исследований для раздражения нервов и мышц применяют электрические стимулы различной формы: прямоугольной, синусоидальной, линейно- и экспоненциально нарастающей, индукционные удары, конденсаторные разряды и т.п. (рис. 2.18).
|
|
|
а |
|
б |
в |
Рис. 2.18. Электрические стимулы различной формы: |
|
|
|
|
а – толчок тока прямоугольной формы; б – линейно |
|||||
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
||
1 |
|
|
|
|
|
нарастающий |
ток; в – экспоненциально нарастающий |
|
|
|
|
|
|
ток; г – |
индукционный замыкательный (1) и |
||
|
|
|
|
|
д |
|
||
|
|
|
|
|
|
размыкательный (2) удары; д – конденсаторный разряд; |
||
2 |
|
|
|
|||||
|
е |
|
е – синусоидальный ток. |
|||||
|
|
|
|
|
|
|||
г
Механизм раздражающего действия тока при всех видах стимулов в принципе одинаков, однако в наиболее отчетливой форме он выявляется при использовании постоянного тока.
36

ФИЗИОЛОГИЯ МЫШЦ |
3 |
|
|
ГЛАВА3. ФИЗИОЛОГИЯМЫШЦ
Морфологически различают три вида мышц /4/:
 поперечно-полосатые скелетные;
поперечно-полосатые скелетные;
 поперечно-полосатая сердечная;
поперечно-полосатая сердечная;
 гладкие внутренних органов, кожи, сосудов.
гладкие внутренних органов, кожи, сосудов.
Функции скелетной мускулатуры /4/:
обеспечение перемещения тела и его отдельных частей в пространстве;
антигравитационные тонические функции, поддержание позы;
обеспечение равновесия тела в различных статических положениях и при движении организма;
участие в терморегуляции (мышцы увеличивают теплопродукцию при сокращении и мышечной дрожи);
сокращение мышц способствует току лимфы;
тоническая «подпитка» функций мозга.
3.1.Физическиеифизиологическиесвойстваскелетных мышц
К физическим свойствам скелетных мыщц относятся:
 растяжимость – способность мышцы изменять длину под действием растягивающей ее силы.
растяжимость – способность мышцы изменять длину под действием растягивающей ее силы.
 эластичность – способность мышцы восстанавливать первоначальную длину после прекращения действия растягивающей силы.
эластичность – способность мышцы восстанавливать первоначальную длину после прекращения действия растягивающей силы.
 сила мышцы – способность мышцы поднять максимальный груз. Для сравнения силы различных мышц определяют их абсолютную силу путем деления максимального поднятого груза на площадь ее физиологического поперечного сечения.
сила мышцы – способность мышцы поднять максимальный груз. Для сравнения силы различных мышц определяют их абсолютную силу путем деления максимального поднятого груза на площадь ее физиологического поперечного сечения.
 способность мышцы совершать работу. Работа мышца определяется произведением величины поднятого груза на высоту мышечного сокращения. Работа мышцы увеличивается с увеличением поднимаемого груза до определенного предела, после которого увеличение груза приводит к снижению высоты подъема и уменьшению работы. Максимальная работа совершается мышцей при перемещении груза средней величины (закон средних нагрузок).
способность мышцы совершать работу. Работа мышца определяется произведением величины поднятого груза на высоту мышечного сокращения. Работа мышцы увеличивается с увеличением поднимаемого груза до определенного предела, после которого увеличение груза приводит к снижению высоты подъема и уменьшению работы. Максимальная работа совершается мышцей при перемещении груза средней величины (закон средних нагрузок).
К физиологическим свойствам скелетных мыщц относятся:
 возбудимость – способность отвечать на действие раздражителя возбуждением.
возбудимость – способность отвечать на действие раздражителя возбуждением.
 проводимость – способность проводить возбуждение от места его возникновения к соседним участкам мышцы.
проводимость – способность проводить возбуждение от места его возникновения к соседним участкам мышцы.
 сократимость – способность мышцы изменять длину или напряжение в ответ на действие раздражителя;
сократимость – способность мышцы изменять длину или напряжение в ответ на действие раздражителя;
 лабильность – способность мышцы сокращаться в соответствии с частотой действия раздражителя. Лабильность скелетных мышц находится в пределах 100-200 имп/с.
лабильность – способность мышцы сокращаться в соответствии с частотой действия раздражителя. Лабильность скелетных мышц находится в пределах 100-200 имп/с.
При непосредственном раздражении мышцы (прямое раздражение) или при раздражении через иннервирующий ее двигательный нерв (непрямое раздражение) одиночным стимулом возникает одиночное мышечное сокращение, состоящее из трех фаз:
 латентного периода, который начинается с момента действия раздражителя и заканчивается с началом фазы укорочения;
латентного периода, который начинается с момента действия раздражителя и заканчивается с началом фазы укорочения;
37

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
 фазы сокращения (укорочения) – от начала укорочения до его максимума;
фазы сокращения (укорочения) – от начала укорочения до его максимума;
 фазы расслабления – от максимума сокращения до восстановления исходной длины
фазы расслабления – от максимума сокращения до восстановления исходной длины
(рис. 3.1) /4/.
|
|
|
1 |
2 |
|
3 |
Рис. 3.1. Одиночное мышечное сокращение (механическая |
|
|
|
|
||||
|
|
|
|
|
|
|
запись): А – сокращению предшествует потенциал действия |
|
|
|
|
|
|
|
(1 – латентный период; 2 – фаза укорочения; 3 – фаза |
|
0,01с |
0,05 сA |
0,06 с |
||||
|
расслабления); Б – изменения возбудимости мышцы. |
||||||
|
|
||||||
100% |
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
Б |
|
|
|
|
|
|
|
|
|
|
|
В естественных условиях к скелетной мышце из ЦНС поступают не одиночные импульсы, а серия импульсов, на которые мышца отвечает длительным сокращением, получившим название тетанического сокращения. Различают два вида тетануса: зубчатый и гладкий. Зубчатый тетанус возникает при такой стимуляции мышцы, когда последующий импульс возбуждения поступает к ней в фазу расслабления (неполная суммация сокращений). Гладкий тетанус в фазу укорочения (полная суммация сокращений) (рис. 3.2).
I |
II |
III |
IV |
V |
Рис. 3.2. Суммация одиночных сокращений поперечно-полосатой мышцы в зависимости от начала 2-го раздражения и образование тетануса: I – одиночные мышечные сокрашения; II, III – зубчатый тетанус; IV – гладкий тетанус (оптимальная частота раздражений);V – пессимум (пессимальная частота раздражений).
Для возникновения суммации необходимо, чтобы интервал между раздражениями имел определенную длительность: он должен быть длиннее рефрактерного периода, иначе на второе раздражение не будет ответа, и короче всей длительности сократительного ответа, чтобы второе раздражение подействовало на мышцу раньше, чем она успеет расслабиться после первого раздражения.
При увеличении частоты раздражения последовательно возникают зубчатый тетанус, гладкий тетанус, оптимум и пессимум. Оптимум возникает при ритмическом раздражении, когда последующее раздражение попадает в фазу повышенной возбудимости – экзальтации предыдущего одиночного сокращения. Пессимум – расслабление мышцы при ее ритмическом раздражении с частотой превышаюшей лабильность ткани, когда последующее раздражение попадает в латентный, особенно в рефрактерный период предыдущего одиночного сокращения. В организме слитное сокращение скелетных мышц достигается последовательным раздражением мышечных волокон. Частота возбуждений, приходящих по соматическим нервам к мышцам, обычно в 10 раз меньше величины лабильности мышцы.
38

ФИЗИОЛОГИЯ МЫШЦ |
3 |
|
|
3.2. Нейромоторные(двигательные) единицы
Нейромоторной единицей называется мотонейрон вместе с группой иннервируемых им мышечных волокон (рис. 3.3) /4/. Причем, количество мышечных волокон в нейромоторной
единице различно. Например, в наружных мышцах глазного яблока их 3-6, в пальцах руки 10-25 (это быстрые моторные единицы), в икроножной мышце ноги более 2000 (это медленные моторные единицы). Чем меньшее количество мышечных волокон иннервируется одним нервным волокном, тем точнее движение. Внутри нейромоторных единиц сокращение отдельных мышечных волокон происходит последовательно в связи с неодновременностью прихода к ним возбуждения, что обеспечивает плавность сокращения мышцы.
У здорового человека в расслабленной мышце электрическая активность в мышце почти отсутствует. При небольшом напряжении мышцы, например, связанном с поддержанием позы, двигательные единиц раздражаются с частотой 5-10 имп/с, при увеличении силы сокраще-
Рис. 3.3. Отдельный мотонейрон иннервирует несколько мышечных волокон – моторная единица.
ния частота повышается до 20-30 имп/с, лишь при максимальном напряжении мышцы она может достигать 50 имп/с. Исследование частоты разрядов двигательных единиц позволило установить, что в естественных условиях сокращения мышц мотонейроны раздражаются со сравнительно низкой частотой.
Каждое отдельное нервное волокно иннервирует несколько мышечных волокон. При этом отдельное мышечное волокно может иннервироваться несколькими нервными волокнами от разных мотонейронов.
Тоническое сокращение скелетных мышц, направленное на сохранение равновесия тела в пространстве, обеспечивают проприорецепторы мышц, нейроны спинного мозга и нервные центры экстрапирамидной системы головного мозга.
Виды сокращений: изометрическое, изотоническое и ауксотоническое. При изометрическом сокращении увеличивается тонус мышцы без изменения ее длины. При изотоническом сокращении мышца укорачивается (концентрическое) или удлиняется (эксцентрическое) без изменения тонуса. При ауксотоническом режиме изменяются напряжение и длина мышцы, что наблюдается при различных видах деятельности.
Сокращения скелетных мышц подразделяются на фазные и тонические. Быстрая (фаз-
ная) активность скелетных мышц обеспечивается белыми мышечными волокнами, которые отличаются значительным содержанием миофибрилл, гликогена и гликолитических ферментов. Гликоген обеспечивает энергетические потребности мышцы. Содержание миоглобина в белых мышечных волокнах незначительно. Белые мышечные волокна, миофибриллы которых толще и сильнее, быстро сокращаются, но и легко утомляются.
Тонические сокращения определяются красными мышечными волокнами, в которых содержится большое количество миоглобина и цитоплазматических органелл, особенно митохондрий. Поэтому красные мышечные волокна способны к длительному напряжению и
39

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
длительной динамической работе. Большинство мышц смешанные, они содержат белые и красные волокна в разных соотношениях.
Быстрые и медленные мышечные волокна имеют разный тип энергетического обеспечения. Белые мышечные волокна, богатые гликогеном, преимущественно работают на энергетической основе анаэробного окисления глюкозы, они быстро утомляются. Медленные красные мышечные волокна, богатые митохондриями и миоглобином, используют энергию аэробных реакций. Они хорошо снабжаются кровью и долго работают без утомления.
3.3. Утомление
Утомление – физиологическое состояние человека, выражающееся во временном снижении работоспособности. Мышечное (физическое) и центральное (нервно-психическое) утомление обычно сочетаются. Утомление характеризуется уменьшением силы и выносливости мышц, нарушением координации движений, ослаблением оперативной памяти, внимания, снижением скорости переработки информации. Причинами утомления могут быть истощение депо гликогена и ослабление процесса ресинтеза АТФ; накопление кислых продуктов метаболизма (фосфорная и молочная кислоты); истощение депо кальция и утомление нервных центров, регулирующих сокращение отдельных групп мышц. Субъективно утомление ощущается в виде усталости и потребности во сне. В эксперименте утомление мышцы развивается вследствие длительного ритмического раздражения. Признаками его являются снижение амплитуды сокращений, увеличение их латентных периодов, удлинение фазы расслабления и прекращение сокращений при продолжающемся раздражении (рис. 3.4) /1/.
Рис. 3.4. Две кривые утомления (эргограммы) студентов, записанные до (а) и после приема (б) в течение 6 ч зачетов (по Моссо).
а |
б |
В целом организме утомляются сначала нервные центры, затем синапсы, и только после этого мышечные волокна.
40