

Нормальная физиология (Пособие для резидентуры)-1
.pdfФИЗИОЛОГИЯ МЫШЦ |
3 |
|
|
3.4. Работаисиламышцы
Согласно законам физики, работа есть энергия, затрачиваемая на перемещение тела с определенной силой на определенное расстояние. Если сокращение мышцы совершается без нагрузки (изотонический режим), то механическая работа равна нулю. Если при максимальной нагрузке не происходит укорочения мышца (изометрический режим), то работа также равна нулю. Согласно закону средних нагрузок, мышца может совершать максимальную работу при нагрузке средней величины.
При сокращении скелетных мышц в естественных условиях, преимущественно в режиме изометрического сокращения, например при фиксированной позе, говорят о статической работе, при совершении движений – о динамической. Статический режим работы более утомителен, чем динамический. Работа может совершаться и в условиях удлинения мышцы (опускание груза), тогда она называется уступающей работой.
Скорость и сила, с которой одна часть тела перемещается относительно другой, зависят от длины рычага, т.е. взаимного расположения точек прикрепления мышц и оси вращения, а также от длины, силы мышцы и величины нагрузки. В зависимости от функции, которую выполняет конкретная мышца, возможно превалирование скоростных или силовых качеств. Причем, чем длиннее мышца, тем больше скорость ее укорочения. При этом большую роль играет параллельное расположение мышечных волокон относительно друг друга. В этом случае физиологическое поперечное сечение, под которым подразумевается сумма поперечных сечений отдельных волокон, входящих в состав данной мышцы, соответствует геометрическому (портняжная мышца) (рис. 3.5). У мышц с перистым расположением мышечных волокон (икроножная мышца) силовые характеристики выше, поскольку физиологическое поперечное сечение больше геометрического. У мышц веретенообразной формы (двуглавая мышца плеча) геометрическое сечение совпадает с физиологическим только в средней части, а в других областях физиологическое сечение больше геометрического, поэтому мышцы этого типа занимают по своим характеристикам промежуточное место. Для сравнения силы разных мышц определяют абсолютную мышечную силу /1, 4/.
Рис. 3.5. Строение различных типов мышц и их физиологические сечения: А – портняжная мышца; Б – икроножная мышца; В – двуглавая мышца плеча /1, 3/.
A Б В
При определении абсолютной силы различных мышц максимальное усилие, которое развивает мышца делят на площадь физиологического поперечного сечения. Например, абсолютная сила икроножной мышцы человека составляет 5,9 кг/см2, двуглавой мышцы плеча – 11,4 кг/см2 /1/.
Для оценки силовых и скоростных характеристик скелетных мышц человека используют метод динамометрии.
41

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
3.5. Механизммышечногосокращения
Каждое мышечное волокно состоит из множества субъединиц – миофибрилл, которые состоят из протофибрилл. Каждая протофибрилла состоит из полимеризованных белковых молекул (миофиламентов) – актина и миозина. Поперечные перегородки – Z-мембраны делят миофибриллы на саркомеры, состоящие из поперечных правильно чередующихся светлых и темных полос. Каждый саркомер состоит из 1500 миозиновых и 3000 актиновых миофиламентов /1/. Миозиновые нити образуют наиболее плотную часть саркомера, называемую А-диском. Будучи анизотропным, он сильно поляризует видимый свет. В центре А-диска проходит светлая полоса, называемая Н-полоской. По обе стороны от А-диска находятся участки, содержащие тонкие нити актина, получившие название I-дисков (изотропных дисков), почти не поляризующих свет. Посередине I-дисков проходит темная линия – Z-мем- брана (рис. 3.6). Благодаря чередованию светлых (I) и темных (А) дисков, скелетные мышцы выглядят поперечно-исчерченными.
A |
I |
A |
I |
A |
|
|
|
а |
|
|
|
|
|
|
|
н |
z |
|
|
Рис. 3.6. |
Структура |
миофибриллы |
|
|
|
(схема). Показаны диски А |
и I, |
||||
|
I |
|
|
||||
|
Н |
|
Z-мембрана и Н-полоска (а). Взаимное |
||||
|
|
|
|
расположение толстых (миозиновых) и |
|||
б |
|
|
|
тонких |
(актиновых) |
нитей |
в |
|
|
|
расслабленной (б) и сокращенной (в) |
||||
|
|
|
|
||||
|
|
|
|
миофибрилле. |
|
|
|
в








































































































Процесс сокращения связан с изменением длины саркомера, что обусловлено перекрытием актиновых и миозиновых филаментов. Во время сокращения А-диски, состоящие в основном из миозина, сохраняют свой размер, в них почти исчезает светлая Н-полоска, так как актиновые нити при сокращении сближаются друг с другом своими концами, диски I, состоящие из актина, укорачиваются. Каждая нить актина образована двумя перекрещивающимися цепями мономеров. В углублениях между цепочками мономеров актина располагаются нити белка тропомиозина, к каждой из которых прикреплен глобулярный белок тропонин. Тропонин состоит из тропомиозинсвязывающих (ТnТ), кальцийсвязывающих (ТnС) и тормозных (ТnI) субъединиц (рис. 3.7) /1, 4/. Тропонин Т обладает сродством к тропомиозину, тропонин С обладает сродством к ионам Са2+, тропонин I – к актину.
42

ФИЗИОЛОГИЯ МЫШЦ |
3 |
|
|
|
Саркомер |
|
|
|
|
Z диск |
|
Миозин |
|
|
|
|
Поперечные |
|
|
||
|
|
мостики |
Актин |
|
|
|
|
|
|
|
|
Толстый |
филамент |
Тонкий |
филамент |
|
|
|
|
|
|
||
Актин |
Миозин |
Актин |
|
Сокращение |
|
Z |
Расслабление |
Z |
|
Z |
Z |
|
|
Головка |
|
|
|
Хвост |
Светлая цепь |
|
|
|
Миозин |
|
134 нм |
|
|
Участок, |
|
|
|
присоединения |
|
Мономеры актина |
|||
|
миозиновой головки |
||
|
|
||
|
|
|
Актиновая спираль
+
Тропомиозин |
I |
С |
|
Т |
Тропонин |
Актиновые
филаменты
Рис. 3.7. Молекулярное строение саркомера.
Теория скольжения нитей, выдвинутая в 1954 году Г.Хаксли и Н.Хэнсоном, исходит из того, что изменение длины саркомера обусловлено взаимным перекрытием актиновых и миозиновых филаментов. При поступлении нервных импульсов к мышце происходит деполяризация мембраны концевой пластинки нервного окончания. ПД по плазматической мембране миофибриллы быстро распространяется по продольным и поперечным Т-трубочкам. Благодаря этому сигнал передается эндоплазматической сети (рис. 3.8). В результате из концевых латеральных цистерн саркоплазматической сети высвобождаются ионы Са2+, которые связываются с тропонином С.
43
НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
2 |
1 |
2 |
1 |
2 |
1 |
2 |
1 |
|
3 |
4 |
3 |
|
3 |
4 |
3 |
|
|
|
|
|
|
||
|
|
|
|
|
|
|
4 |
4 |
|
5
5
Z |
Z |
Z |
Z |
А |
|
|
Б |
Рис. 3.8. Схематическое изображение взаимоотношений поверхностной мембраны (1), поперечных трубочек (2), боковых цистерн (3) и продольных трубочек (4) саркоплазматического ретикулума и миофибрилл (5) мышечного волокна. Z-мембрана: А – в состоянии покоя, Б – во время сокращения. Деполяризация мембраны и поперечных трубочек вызвала освобождение ионов Са2+ из боковых цистерн. Освобождавшийся Са2+ диффундирует по направлению к фибриллам и частично захватывается продольными трубочками ретикулума.
В исходном состоянии взаимодействие между актином и миозином тормозится субъединицей тропонина I. В результате связывания ионов Са2+ с субъединицей тропонина С тормозящее влияние тропонина I на актин ослабевает и тропомиозин смещается латерально
(рис. 3.9) /1/.
Состояние |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Актин |
|||||||||||
|
|
|
|
|
|
|
Тропомиозин |
||||||||||||||||||
|
|
покоя |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
Тонкий филамент |
|
|
|
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Тропонин |
|||
|
|
|
Вид поперечного |
|
|
|
|
|
|
|
|||||||||||||||
|
|
|
|
Участок связывания с |
|
||||||||||||||||||||
|
|
|
|
|
Поперечный |
||||||||||||||||||||
|
|
|
|
|
разреза |
|
миозином (миозина) |
|
|||||||||||||||||
|
|
|
|
|
|
|
|
мостик |
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
Участок связывания |
|
|
|
|
|
|
|||||||
|
|
Актин |
|
|
|
|
|
|
|
с миозином |
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
Тропо- |
Тропомиозин |
|
|
|
|
|
|
|||||||||||||||||
|
|
нин |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
Участок связывания |
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
миозина с актином |
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
При связывании Са2+ |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
Поперечный мостик |
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
тропонином С, тропо- |
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
миозин смещается и тем |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
самым создаются условия |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
для взаимодействия |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
актина с миозином |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
При |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Са2+ |
Са2+ |
сокращении |
|
|||||||
|
|
|
|
|
Са2+ |
|
|
|
|
|
|
|
|
||||||||||||
Рис. 3.9. Механизм мышечного сокращения.
Это в свою очередь освобождает связывающие места – активные центры для прикрепления миозиновой головки. Последняя, прикрепляясь к связывающий зоне молекулы актина,
44

ФИЗИОЛОГИЯ МЫШЦ |
3 |
|
|
поворачивается под углом в 90°, вызывая скольжение нити актина. В головках миозина содержится фермент аденозинтрифосфатаза (АТФ-аза), который при контакте головки миозина с актином осуществляет гидролиз фосфатных связей АТФ, после чего происходит отсоединение головки миозина и последующее ее прикрепление к следующему месту связывания актина. Каждая толстая нить миозина имеет около 500 головок, и каждая за время быстрого сокращения мышцы осуществляет около пяти раз циклы взаимодействия с актином.
Организация скелетной мышцы от макроскопического до молекулярного уровня показана на рис. 3.10 /7/.
СКЕЛЕТНАЯ МЫШЦА
Мышца
Пучок мышечных волокон
Z-мембрана |
Мышечное волокно |
|
|||
Н-полоска |
|
А-диск I-диск |
|
|
|
|
|
Z СаркомерZ |
Миофибрилла |
Молекулы G-актина |
|
|
|
Н |
|
||
|
|
|
|
|
|
|
|
|
Миофиламенты |
|
|
|
|
|
|
Нить F-актина |
|
|
|
|
|
Нить миозина |
|
|
|
|
|
Молекула миозина |
|
|
|
|
|
Легкий |
Тяжелый |
Е |
|
|
|
меромиозин |
|
Ж |
З |
И |
меромиозин |
||
Рис. 3.10. Организация скелетной мышцы от макроскопического до молекулярного уровня. Е, Ж, З, И – поперечное сечение на указанном уровне /7/.
При отсутствии повторного импульса возбуждения концентрация ионов кальция в протофибриллярном пространстве уменьшается за счет кальциевого насоса, обеспечивающего реаккумуляцию ионов Са2+ в систему цистерн саркоплазматического ретикулума, где они хранятся до следующих процессов возбуждения.
Энергообеспечение работы кальциевого насоса происходит за счет энергии, образующейся при гидролизе АТФ. АТФ является важным фактором, необходимым и для процесса расслабления.
Таким образом, в процессе сокращения мышечного волокна в нем происходят следующие преобразования:
 генерация ПД;
генерация ПД;  распространение ПД по Т-системе;
распространение ПД по Т-системе;
45

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
 электрическая стимуляция зоны контакта Т-трубочек и саркоплазматического ретикулума, повышение внутриклеточной концентрации ионов Са2+;
электрическая стимуляция зоны контакта Т-трубочек и саркоплазматического ретикулума, повышение внутриклеточной концентрации ионов Са2+;
 взаимодействие ионов Са2+ с тропонином С, освобождение активных центров на актиновых филаментах;
взаимодействие ионов Са2+ с тропонином С, освобождение активных центров на актиновых филаментах;
 взаимодействие миозиновой головки с актином, вращение головки и развитие эластической тяги;
взаимодействие миозиновой головки с актином, вращение головки и развитие эластической тяги;  скольжение актина и миозина относительно друг друга, уменьшение размера сарко-
скольжение актина и миозина относительно друг друга, уменьшение размера сарко-
мера, развитие напряжения или укорочения мышечного волокна.
Энергетикамышечногосокращения
В динамическом режиме работоспособность мышцы определяется скоростью расщепления и ресинтеза АТФ. Ресинтез АТФ может обеспечиваться за счет окислительного расщепления глюкозы. Действительно, при умеренных нагрузках ресинтез АТФ обусловлен повышенным потреблением мышцами глюкозы и кислорода. У тренированных людей большую роль в обеспечении повышенной потребности организма в энергии играет повышение активности митохондриальных ферментов.
При максимальной физической нагрузке происходит дополнительное расщепление глюкозы путем анаэробного гликолиза. Во время этих процессов ресинтез АТФ осуществляется в несколько раз быстрее, и механическая работа, производимая мышцами, также больше, чем при аэробном окислении. Предельное время для такого рода работы, составляет около 30 с, после чего возникает накопление молочной кислоты, т.е. метаболический ацидоз, и развивается утомление.
Таким образом, химический ресинтез АТФ осуществляется в работающих мышцах анаэробным (без участия кислорода) и аэробным (с участием кислорода) путем. Быстрые и медленные мышечные волокна имеют разный тип энергетического обеспечения. Быстрые мышечные волокна, богатые гликогеном, работают преимущественно на энергетической основе анаэробной фазы мышечного сокращения, они быстро утомляются. Медленные мышечные волокна, богатые митохондриями и миоглобином, при работе используют в основном энергию аэробных реакций. Они хорошо кровоснабжаются и могут долго работать без утомления /4/.
Нарушение ресинтеза АТФ ядами, подавляющими гликолитические и окислительные процессы, ведет к полному исчезновению АТФ и креатинфосфата, вследствие чего кальциевый насос перестает работать. Концентрация Са2+ в области миофибрилл значительно возрастает и мышца приходит в состояние длительного необратимого укорочения.
Законсреднихнагрузок
Сила сокращения зависит от степени растяжения мышцы перед ее сокращением. Наибольшая амплитуда сокращения достигается при средней нагрузке, при этом между нитями актина и миозина возникает максимальное количество поперечных мостиков (рис. 3.11Б). При сильной нагрузке длина саркомеров увеличивается, и количество поперечных мостиков между нитями актина и миозина уменьшается, что приводит к меньшей силе сокращения (рис. 11В). При отсутствии нагрузки количество поперечных мостиков между
46

ФИЗИОЛОГИЯ МЫШЦ |
3 |
|
|
актином и миозином также уменьшается, поскольку нити актина приближаются друг к другу при уменьшении длины саркомера (рис. 3.11А) /4/.
|
|
а |
|
Ас |
б |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
А |
Р |
|
|
|
|
|
|
Р |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Б |
|
|
|
|
|
|
|
|
|
|
|
|
|
Ак Ак
М
Р
В |
t |
Рис. 3.11. Одиночные сокращения скелетной мышцы при предварительном растяжении ее разной силой: А – при сокращении мышцы без нагрузки сила мышечного сокращения ниже максимальной, так как нити актина мешают друг другу контактировать с миозином; Б – при средней нагрузке максимальная сила сокращения обусловлена максимальным количеством поперечных мостиков между актином и миозином; В – при максимальной нагрузке сила мышечного сокращения уменьшается за счет снижения количества поперечных мостиков между актином и миозином; а – количество контактов между актином и миозином при изменении длины саркомера; б – амплитуда сокращения мышцы при изменении нагрузки на мышцу (Р – раздражение, Ас – амплитуда сокращения; t – время; Ак – актин; М – миозин).
Мощность мышцы, измеряемая величиной работы в единицу времени, также достигает максимальной величины при средних нагрузках.
Регуляциятонусамышц
Мышечный тонус – состояние продолжительного поддерживания определенного уровня активности мышцы. Тонус мышцы определяется активностью отдельных экстрафузальных мышечных волокон при поступлении к ним импульсов по двигательным волокнам от α-мото- нейронов спинного мозга.
Поддержание мышечного тонуса достигается преимущественной активностью красных мышц, содержащих большое количество миоглобина и имеющих низкую скорость распространения возбуждения и развития сокращения. У человека таких мышц больше среди разгибателей, создающих вертикальное положение тела и работающих против гравитационных сил.
47

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
В формировании мышечного тонуса принимают участие рецепторы мышечных веретен, сухожильных органов, а также рецепторы, расположенные в суставной оболочке.
Мышечные веретена состоят из соединительнотканной капсулы с интрафузальными мышечными волокнами, к которым подходят аксоны γ-мотонейронов (рис. 3.12). В капсуле веретена имеются чувствительные окончания двух типов. Одни возбуждаются при статических, а другие – при динамических, растягивающих веретено нагрузках. Эти чувствительные окончания относятся к рецепторам растяжения. Чем больше в мышце общее количество мышечных веретен, тем тоньше и точнее осуществляемые движения. Мышечные веретена крепятся одним своим концом к мышечному сухожилию, а другим – к собственным фасциям мышцы, т.е. параллельно рабочим (экстрафузальным) мышечным волокнам.
|
|
Аксон γ-эфферента |
|
|
спинного мозга |
|
Мотонейронный |
|
|
пул |
|
α-Мотонейрон |
Капсула |
γ-Афферентное |
|
||
|
|
волокно |
|
|
Чувствительные |
|
|
окончания |
Моторная |
|
Интрафузальные |
|
мышечные волокна |
|
единица |
|
|
|
|
Рис. 3.12. Моторные элементы сегментов спинного мозга и мышечное веретено /4/.
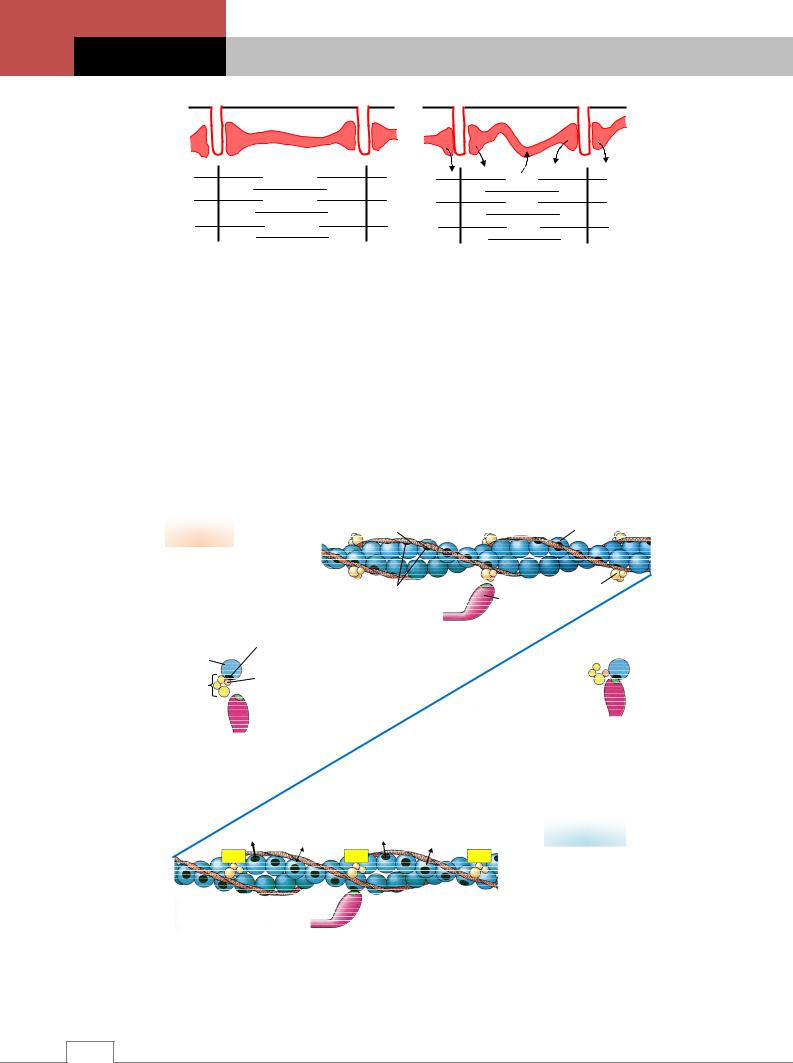
Саморегуляция мышечного тонуса. Мышечный тонус поддерживается на основе принципа саморегуляции: изменение мышечного тонуса вызывает процессы, направленные на его восстановление. Саморегуляция мышечного тонуса осуществляется сегментарным аппаратом спинного мозга. Тоническая активность нейронов спинного мозга определяется как импульсацией, поступающей к ним от кожных рецепторов и проприорецепторов мышц, так и нисходящими центральными влияниями. Перерезка задних корешков спинного мозга у спинальной лягушки на одной стороне приводит к полному расслаблению мышц соответствующей лапки и ее провисанию (опыт Бронджеста) /4/.
При устранении нисходящих влияний (травматический разрыв спинного мозга или его местная анестезия) также наблюдается выраженное снижение тонуса мышц.
Роль проприорецепторов. В сегментарном аппарате спинного мозга наряду с α-мото- нейронами, возбуждение которых вызывает сокращение экстрафузальных мышечных волокон, имеется группа γ-мотонейронов, посылающая импульсы к собственным рецепторам мышцы – мышечным веретенам (рис. 3.13) /4/.
48
|
ФИЗИОЛОГИЯ МЫШЦ |
3 |
Мозжечок |
Красное ядро |
|
Вестибулярное |
Ретикулярная формация |
|
ядро |
|
|
среднего мозга |
|
|
|
|
|
|
Сухожильный |
|
Лабиринт |
орган Гольджи |
|
γ -афферент |
|
|
|
Мышца |
|
Тормозной |
|
|
нейрон |
γ-эфферент |
|
|
|
|
α-мотонейрон |
γ-мотонейрон |
|
Мышечное |
|
|
|
|
|
|
веретено |
|
α-эфферент
Рис. 3.13. Взаимоотношение рецепторов мышцы с моторными элементами сегмента спинного мозга (схема) /4/.
Роль сухожильных рецепторов. Кроме мышечных веретен в саморегуляции мышечного тонуса принимают участие рецепторы, расположенные в сухожилиях мышц (рецепторы Гольджи). Возбуждение сухожильных рецепторов происходит как при пассивном растяжении, так и при активном сокращении мышц. Возбуждение, идущее от сухожильных рецепторов по γ-афферентным волокнам, поступает к α-мотонейронам соответствующих сегментов спинного мозга, которые иннервируют эти мышцы и вызывает (при участии тормозных клеток Реншоу) их торможение. Часть импульсов от сухожильных рецепторов распространяется и к α-мотонейронам мышц-антагонистов /1, 4/.
Рецепторы суставов. К рецепторам, участвующим в поддержании мышечного тонуса,
относятся рецепторы суставов, которые возбуж- |
|
даются при растяжении суставных сумок. Уровень |
|
активации этих рецепторов соответствует углу |
|
поворота конечности в суставе. Диапазон углов сме- |
|
щения, при котором возникает возбуждение сустав- |
|
ных рецепторов, колеблется от 2-3° до 30°. |
А |
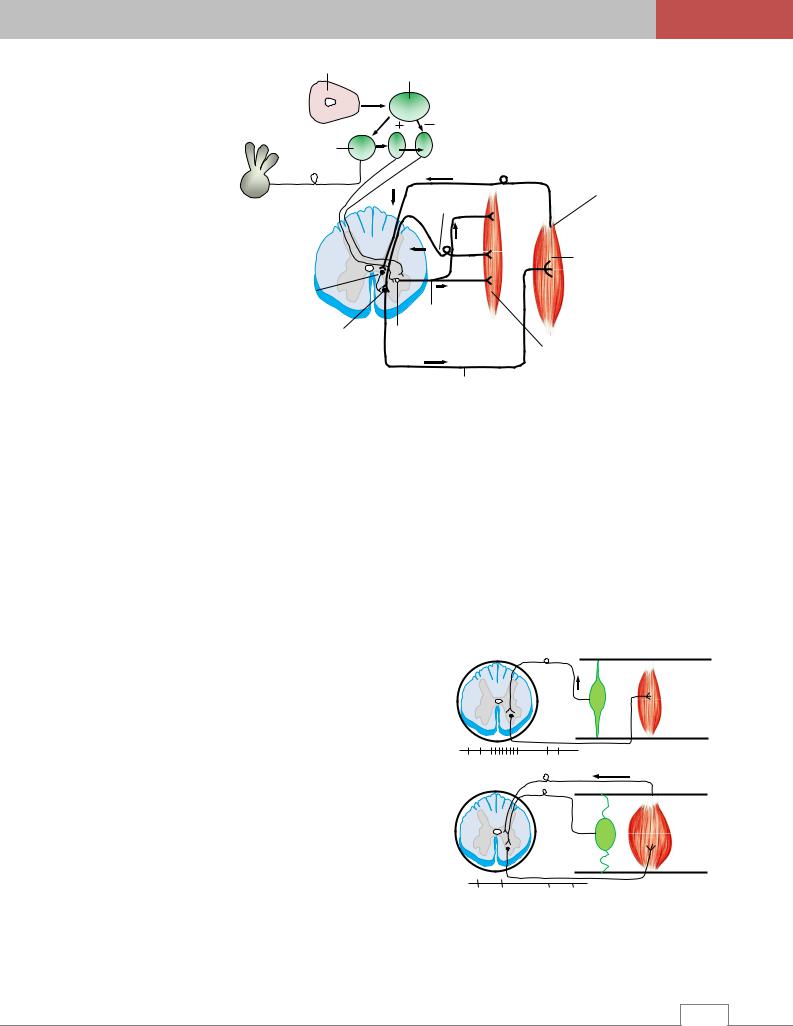
Механизм саморегуляции мышечного тону- |
|
са. При пассивном растяжении мышцы возбужде- |
|
ния от чувствительных окончаний γ-афферентных |
|
волокон мышечного веретена через задние корешки |
|
поступают в спинной мозг и переключаются на |
|
α-мотонейроны передних рогов спинного мозга того |
Б |
же сегмента. Это приводит к повышению возбуди- |
Рис. 3.14. Импульсация от мышечных |
мости α-мотонейронов и последующему усилению |
|
тонуса мышцы (рис. 3.14). При сокращении мышцы |
веретен при расслаблении (А) и |
чувствительные окончания веретена перестают воз- |
сокращении (Б) скелетной мышцы /4/. |
|

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
буждаться, вследствие чего прекращается поток импульсов, идущих к α-мотонейронам, уровень их возбудимости падает и тонус мышцы снижается. С помощью этого механизма мышечные веретена воспринимают длину мышцы и скорость ее изменения /4/.
Активация α-мотонейронов дополнительно приводит к сокращению экстрафузальных волокон и увеличению объема капсулы веретена.
Координация процессов саморегуляции мышечного тонуса на уровне сегментов спинного мозга достигается также возвратным торможением разрядной деятельности мотонейронов при участии тормозных клеток Реншоу. Активация самих клеток Реншоу происходит под влиянием импульсаций, поступающих к ним от γ-афферентов мышечных веретен /4/.
3.6. Физиологическиеособенностигладкихмышц
Гладкие мышцы находятся в стенках внутренних органов, кровеносных и лимфатических сосудов, в коже и морфологически отличаются от скелетной и сердечной мышц отсутствием видимой поперечной исчерченности.
Гладкие мышцы подразделяются на висцеральные (унитарные) и мультиунитарные. Унитарные гладкие мышцы находятся во всех внутренних органах, протоках пищеварительных желез, тонких кровеносных и лимфатических сосудах, коже, матке, мочеточниках. К мультиунитарным относятся ресничная мышца, мышцы радужки глаза, мигательной перепонки, крупных кровеносных сосудов, которые функционируют изолированно, независимо друг от друга, иннервируются отдельными нервными окончаниями. В унитарных гладких мышцах благодаря плотным контактам между соседними миоцитами – нексусам потенциалы действия распространяются с одной мышечной клетки на другую, в результате висцеральные (унитарные) гладкие мышцы сокращаются одномоментно с приходом нервного импульса и представляют собой функциональный синцитий.
Особенностью гладких мышц является их способность осуществлять относительно медленные ритмические и длительные тонические сокращения. Относительно медленные, ритмические сокращения гладких мышц желудка, кишечника, мочеточников и других полых органов обеспечивают перемещение их содержимого. Длительные тонические же сокращения гладких мышц сфинктеров полых органов препятствуют свободному выходу из этих органов их содержимого. Гладкие мышцы стенок кровеносных сосудов, особенно артерий и артериол также находятся в состоянии постоянного тонического сокращения. Изменение тонуса мышц стенок артериальных сосудов влияет на уровень артериального давления и кровоснабжение органов.
Функцииисвойствагладкихмышц
Электрическая активность. Висцеральные (унитарные) гладкие мышцы характеризуются нестабильным мембранным потенциалом. Колебания мембранного потенциала независимо от нервных влияний вызывают нерегулярные сокращения, которые поддерживают мышцу в состоянии постоянного частичного сокращения – тонуса. Тонус гладких мышц выражен в сфинктерах полых органов: желчном, мочевом пузырях, в месте перехода желудка в двенадцатиперстную кишку и тонкой кишки в толстую, а также в гладких мышцах
50