1.Процессы образования конечных продуктов обмена простых белков. Основные источники аммиака. Роль глутамина в оезвреживании аммиака и синтезе ряда соединений(как донор амидной группы).
Источники аммиака:
1)дезаминирование АК(в тканях и кишечнике)
2)дезаминирование аминов
3)дезаминирование азотистых оснований
Аммиак в крови – 12-65мкмоль/л(10-120мкг%), в моче – 35,7 – 71,4ммоль/сут(0,5-1,0г)
Аммиак исключительно токсичен.
Обезвреживание:
1)образование амидов(локально)
Гутамат + NH3,NH4+,АТФ, магний++, глутамин-синтетазаглутамин +АДФ +Фн
Глутаминпочки( –аммиак, глутаминаза) Глутамат -аммиак2аммоний+аммониогенез
альфа-КГ
печень, синтез мочевины
синтез пуринов, пиримидинов.
2)восстановительное аминирование
А. альфа-КГ (глутаматДГ, аммоний, 2Н, НАДФ)глутамат, Н2О, НАДФН
Б. глутамат + ПВК (трансаминирование)альфа-КГ +ала
3)образование аммонийных солей
4)синтез мочевины.
2.Депонирование и мобилизация жиров в жировой ткани, физиологическое значение. Транспорт и использование жрных кислот, образующихся при мобилизации жиров. Биосинтез и использование кетоновых тел.
3.Витамин рр. Химическая природа. Растпространение, участие в обменных процессах.
Никотиновая к-та/ниацин/никотинамид/антипеллагрический витамин. Водорастворимый.
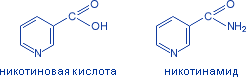 Потребность
18мг. Рис, хлеб, картофель, мясо, печень,
почки, морковь. Актив.форма – НАД, НАДФ.
Потребность
18мг. Рис, хлеб, картофель, мясо, печень,
почки, морковь. Актив.форма – НАД, НАДФ.
Вход.в состав НАД, НАДФ – коферменты дегидрогеназ в ОВР. Процессы биологического окисления(промежуточные переносчики электронов и протонов)
Авитаминоз: пеллагра – дерматиты, диарея, деменция, анорексия, тошнота, понос, обезвоживание. Стоматиты, гингивиты. Головные боли, депрессии, психозы.
4.Способы определения белка в моче.
1.Турбидиметрические методы. основаны на преципитации белка различными агентами: сульфосалициловой кислотой (ССК), трихлоруксусной кислотой (ТХУ), бензетоний хлоридом.
2. Колориметрические методы. К группе колориметрических методов определения белка относятся методы Лоури, биуретовый и методы, основанные на связывании белка с органическими красителями.
3. Определение белка диагностическими полосками. Диагностические полоски позволяют быстро провести полуколичественную оценку содержания белка в моче. По интенсивности сине-зеленой окраски реакционной зоны, развивающейся после кратковременного контакта полоски с мочой, содержащей белок, судят о содержании белка в моче. Основной предпосылкой получения надежных результатов является поддержание стабильного рН, обеспечивающего связывание белком индикатора в стандартных условиях.
4. Определение белка в моче в системе внешней оценки качества
Билет№25
1. Распад пуриновых и пиримидиновых азотистых оснований. Конечные продукты. Пути выведения.
2. Желчные кислоты, строение. Свойства. Участие в переваривании и всасывании липидов. Конъюгация желчных кислот, биологическая роль.
Желчные к-ты – конечный продукт обмена ХС – производные холановой к-ты:
Желчные кислоты:
- первичные 3,7,12-триоксихолановая = холевая; 3,7-диоксиходановая = хенодезоксихолевая(образ.из печени из ХС)
- вторичные 3,12-диоксихолановая = дезоксихолевая; 3-оксихолановая=литохолевая(образ.в кишечнике под действием м/флоры)
**Конъюгирование желчных кислот: 1)с глицином – при у/в питании; 2)с таурином(образ.из цистеина)-при Б питании

роль: - эмульгируют жиры; -активируют липазу, -способсвуют всасыванию продуктов гидролиза жиров в кишечнике.
В сутки из ХС образ. 1г желчи в гепатоцитах→по печеночным пузырным протокам→концентр.в желч.пузыре.Опорожнение происходит постоянно, но усил.под действием поступившей пищи и ХЦкинина.
После выполенния своих ф.85%желчных кислот всас.в подвздошной кишке и вновь поступ.в состав секретируемой желчи – энтерогепатальный круг.
Общий пул желч.к-т 2,8-3,5г. Совершают 6-8 оборотов в сут.
Недостаток желч.к-т: желчекам.болезнь, стеаторрея.
Витамин В6. Химическая природа, распространение, участие в обменных процессах.
Антидерматитный. Адермин, пиридоксин. Водрастворимый.
Прозводные 3-оксипиридина:
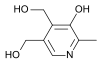 пиридоксин
пиридоксин
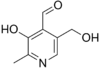 пиридоксаль
пиридоксаль
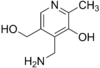 пиридоксамин
пиридоксамин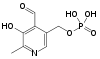 акт.форма-
пиридоксальфосфат.
акт.форма-
пиридоксальфосфат.
Сут.потр. 2мг. Хлеб, горох, фасоль, картофель, мясо, почки, печень.
Коферментные ф.выполняют только фосфорилированные производные – ПФ. 20пиридоксалевых ферментов, катализирующих ключевые р.азотистого метаболизма в организме.
Авитаминоз:акродилия – дерматит с преим.поражением кожи лопаток, носа, ушей. Эпилептиформные припадки; дегенерация ЦНС. Гомоцистинурия, цистатионурия, нарушения обмена трп.
Глюкозурия и ее причины.
Обычно присутствие глюкозы в моче (глюкозурия) является результатом нарушения углеводного обмена вследствие патологических изменений в поджелудочной железе (сахарный диабет, острый панкреатит и т.д.). Реже встречается глюкозурия почечного происхождения, связанная с недостаточностью резорбции глюкозы в почечных канальцах. Как временное явление глюкозурия может возникнуть при некоторых острых инфекционных и нервных заболеваниях, после приступов эпилепсии, сотрясения мозга.
Отравления морфином, стрихнином, хлороформом, фосфором также обычно сопровождаются глюкозурией. Наконец, необходимо помнить о глюкозурии алиментарного происхождения, глюкозурии беременных и глюкозурии при нервных стрессовых состояниях (эмоциональная глюкозурия).
Билет№26
Окисление высших жирных кислот. Последовательность реакций бета-окисления. Связь окисления жирных кислот с цитратным циклом и дыхательной цепью.
Процесс окисления жирных кислот складывается из следующих основных этапов.
Активация жирных
кислот.
Свободная жирная
кислота независимо
от длины углеводородной цепи является
метаболически инертной и не может
подвергаться никаким биохимическим
превращениям, в том числе окислению,
пока не будет активирована. Активация жирной
кислоты протекает
на наружной поверхности мембраны митохондрий при
участии АТФ, коэнзима A
(HS-KoA) и ионов Mg2+. Реакция
катализируется ферментом ацил-КоА-синтетазой: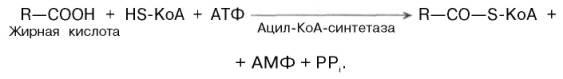
В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты.
Первая стадия дегидрирования. Ацил-КоА в митохондриях прежде всего подвергается ферментативному дегидрированию, при этом ацил-КоА теряет 2 атома водорода в α- и β-положениях, превращаясь в КоА-эфир ненасыщенной кислоты.
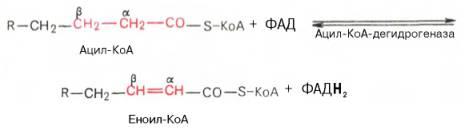
Стадия гидратации. Ненасыщенный ацил-КоА (еноил-КоА) при участии фермента еноил-КоА-гидратазы присоединяет молекулу воды. В результате образуется β-оксиацил-КоА (или 3-гидроксиацил-КоА):
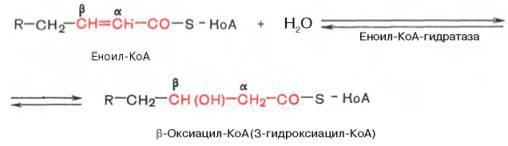
Вторая стадия дегидрирования. Образовавшийся β-оксиацил-КоА (3-гидроксиацил-КоА) затем дегидрируется. Эту реакциюкатализируют НАД+-зависимые дегидрогеназы:
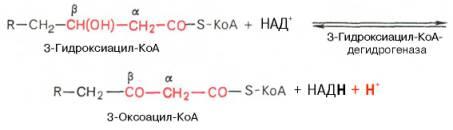
Тиолазная реакция. представляет собой расщепление 3-оксоацил-КоА с помощью тиоловой группы второй молекулы КоА. В результате образуется укороченный на два углеродных атома ацил-КоА и двууглеродный фрагмент в виде ацетил-КоА. Данная реакция катализируется ацетил-КоА-ацилтрансферазой (β-ке-тотиолазой):
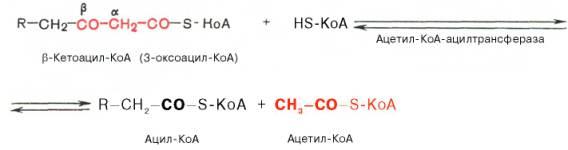
Образовавшийся ацетил-КоА подвергается окислению в цикле трикар-боновых кислот, а ацил-КоА, укоротившийся на два углеродных атома, снова многократно проходит весь путь β-окисления вплоть до образования бутирил-КоА (4-углеродное соединение), который в свою очередь окисляется до 2 молекул ацетил-КоА.
Баланс энергии. При каждом цикле β-окисления образуются одна молекула ФАДН2 и одна молекула НАДН. Последние в процессеокисления в дыхательной цепи и сопряженного с ним фосфорилирования дают: ФАДН2 – 2 молекулы АТФ и НАДН – 3 молекулы АТФ, т.е. в сумме за один цикл образуется 5 молекул АТФ. При окислении пальмитиновой кислоты образуется 5 х 7 = 35 молекул АТФ. В процессе β-окисления пальмитиновой кислоты образуется 8 молекул ацетил-КоА, каждая из которых, «сгорая» в цикле трикарбоновых кислот, дает 12 молекул АТФ, а 8 молекул ацетил-КоА дадут 12 х 8 = 96 молекул АТФ.
Таким образом, всего при полном β-окислении пальмитиновой кислоты образуется 35 + 96 = 131 молекула АТФ. С учетом одноймолекулы АТФ, потраченной в самом начале на образование активной формы пальмитиновой кислоты (пальмитоил-КоА), общий энергетический выход при полном окислении одной молекулы пальмитиновой кислоты в условиях животного организма составит 131 – 1 = 130 молекул АТФ.
Витамин А. Химическая природа, распространение, участие в обменных процессах.
Жирорастворимый. Антиксерофтальмический/ретинол. Депонируется в печени. Сут.потребность 1-2,5мг.
Только животные продукты:печень, яич.желток, молочные продукты, рыбий жир. В растит.продуктах каротиноиды – провитамины А. Актив.форма – ретиналь.
Регуляция роста и дифференцировки клеток(ретино-ретиналь-ретиноевая к-та);
участие в зрит.акте. Палочки: сумуречное зр.(пигмент родопсин);колбочки:цвет.зр.(йодопсин). Кофермент – 11-цис-ретиналь, альд.произв вит.А.
Антиоксидант.
Авитаминоз: гемералопия(сумеречная слепота), ксерофтальмия – сухость роговицы глаза(закупорка слезканала – ороговение эпителия), конъюнктивит, отек, изъязвление роговицы – кератомаляция. Избыточное ороговение кожи, поражения эптелия ЖКТ, мочепол.с-мы, дыхат.с-мы.
Соединительная ткань. Классификация. Клеточные элементы. Основные белки соединительной ткани. Межклеточный матрикс, представление о гликопротеинах соединительной ткани.
1)собственно СТк
2)скелет СТк(хрящевая, костная. в т.ч. зубы)
3)СТк со спец.св-вами: жировая, слизистая, пигментная, ретикулярная, кровь, лимфа, синовиальная и т.д.
Общие свойства: происхождение из мезенхимы, сходство стр-ры, выполняют опорные функции.
СТк – межкл.матрикс с клетками различно типа и волокнистыми стуктурами.
По физиологич.состоянию: волокнистые, твердые(кости), гелеобразные(хрящи), жидкие.
Собственно СТк распределена: по ходу сосудов, в обл.почечных лоханок, вход.в сост.хрящей связок сухожилий, сост.основу м/кл.в-ва паренхиматозных органов, подстил.кожу.
Коллаген.
основной Б СТк. Составляет 25-33% от общего кол-ва Б, 6% от массы тела. Химический состав: 1/3 АК глицин, 1/5 пролин, оксипролин, 1/10 аланин.
Генетические дефекты коллагена:
дерматоспораксис – заболев.круп.рогат.скота – повышенная ломкость кожи. Дефект проколлагеновой пептидазы.
латиризм – ингибирование лизиноксидазы, аминооксидазы
синдром Элерса-Данлоса: коллагенозы
несовершенный остеогенез(врожд.ломкость костей, б.»стеклянного мужчины», врожд.рахит)
синдром Марфана
врожденный атеросклероз. Коллаген 3 типа в сосудах.
50% коллагена обновляется за 50 лет.
Эластин.
Беден о-пролином, о-лизином. 1/3 часть АК – глицин. Основа стр-ры: десмозин, изодесмозин.
Десмозин.
Обеспечивает растяжимоть, стретч-свойства. Образ-ся из 4 остатков лизина. Нарушение резиноподобных св-в приводит: эмфизема, аневризмы сосудов. Причины: нарушение активности эластазы при дефиците меди и витамина В6, разрушение эластина под действием эластазы. Эластазу синт.нейтрофилы(в легких – эмфизема!). В норме не происходит, т.к. эластазу ингибирует альфа-1-антитрипсин(б.острой фазы), синтезируемый альвеолярными макрофагами, что защищает от действия эластазы. При дефиците – усиленное действие эластазы – эмфизема.
Основное в-во СТк. Межклеточный матрикс.
Представлено гликопротеинами:
1)собственно ГП(95%белка и 5%у/в нерегулярного строения) – муцины, гаптолобины, ФСГ, ЛГ и др.
2)протеогликаны(5%белок и 95%ГАГ) Небелковая часть: ГАГ – ВМС, ВПС(кислые мукополисахариды). Мономер – дисахаридная единца(у/в регулярного строения)
Заболевния: коллагенозы, остеопороз, врожд.пороки сердца, сколиозы, близорукость, гипермобильность суставов, дисплазия СТк, опущение органов.
Билет№27
Обмен триптофана. Образование серотонина, биологическая роль. Кинурениновый и серотониновый пути превращения триптофана.
Трп – незаменимая АК. В физиологических условиях более 95% триптофана окисляется по кинурениновому пути и не более 1% – по серотониновому. Серотонин в организме подвергается окислительному дезаминированию с образованием индолилук-сусной кислоты, которая выделяется с мочой. Содержание этой кислоты в моче повышено при поражениях кишечника злокачественными карци-ноидами, когда около 60% триптофана окисляется по серотониновому пути.
серия р-ций→серия р-ций +ФРПФ→рибонуклеотид никотин.к-ты +АТФ, +глутамин→НАД+ перенос е в дых.цепи→НАДФ+
ТРп гидроксилаза, О2→Н2О →5-окситриптофан декарбоксилаза, ПФ→ серотонин
- сужает сосуды
- стимулирует активность мозга
- регулирует сон, поведение, кровяное давление, температуру
- стимулирует секрецию гастроинтестинальных Г
- стимулирунт перистальтику
- серотонин – мелатонин(образ.в шишковид.железе)
Биосинтез жирных кислот, последовательность реакций. Регуляция биосинтеза.
Синтез жирных кислот протекает в цитоплазме клетки. В митохондриях в основном происходит удлинение существующих цепей жирных кислот. Установлено, что в цитоплазме печеночных клеток синтезируется пальмитиновая кислота (16 углеродных атомов), а в митохондриях этих клеток из уже синтезированной в цитоплазме клетки пальмитиновой кислоты или из жирных кислот экзогенного происхождения, т.е. поступающих из кишечника, образуются жирные кислоты, содержащие 18, 20 и 22 углеродных атома.
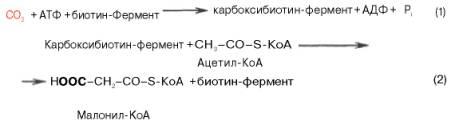
Первой реакцией биосинтеза жирных кислот является карбоксилирование ацетил-КоА, для чего требуются бикарбонат, АТФ, ионы марганца. Катализирует эту реакцию фермент ацетил-КоА-кар-боксилаза. Фермент содержит в качестве простетической группы биотин. Авидин – ингибитор биотина угнетает эту реакцию, как и синтез жирных кислот в целом.
Установлено, что ацетил-КоА-карбоксилаза состоит из переменного числа одинаковых субъединиц, каждая из которых содержит биотин, биотинкарбоксилазу, карбоксибиотинпереносящий белок, транскарбоксилазу, а также регуляторный ал-лостерический центр, т.е. представляет собой полиферментный комплекс.
Реакция протекает в два этапа: I – карбоксилирование биотина с участием АТФ и II – перенос карбоксильной группы на ацетил-КоА, в результате чего образуется малонил-КоА:
Мультиферментный комплекс, называемый синтетазой (синтазой) жирных кислот, состоит из 6 ферментов, связанных с так называемым ацилпереносящим белком (АПБ). Данный белок в синтетазной системе выполняет роль КоА.Приводим последовательность реакций, происходящих при синтезе жирных кислот:
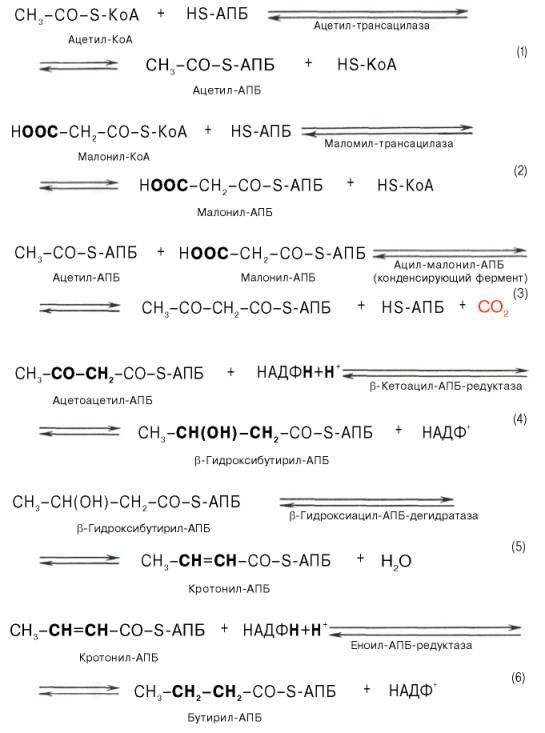 образованием
бутирил-АПБ завершается лишь первый из
7 циклов, в каждом из которых началом
является присоединение молекулы
малонил-АПБ к карбоксильному концу
растущей цепи жирной кислоты. При этом
отщепляется дистальная карбоксильная
группа малонил-АПБ в виде СО2. Например,
образовавшийся в первом цикле бутирил-АПБ
взаимодействует с малонил-АПБ:
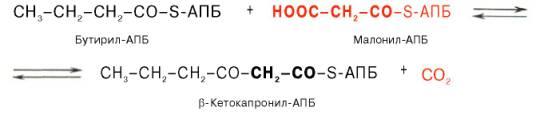
образованием
бутирил-АПБ завершается лишь первый из
7 циклов, в каждом из которых началом
является присоединение молекулы
малонил-АПБ к карбоксильному концу
растущей цепи жирной кислоты. При этом
отщепляется дистальная карбоксильная
группа малонил-АПБ в виде СО2. Например,
образовавшийся в первом цикле бутирил-АПБ
взаимодействует с малонил-АПБ:
Завершается синтез
жирной кислоты отщеплением HS-АПБ от
ацил-АПБ под влиянием фермента деацилазы.
Например:
![]()
Суммарное уравнение
синтеза пальмитиновой кислоты можно
записать так:

Образование ненасыщенных жирных кислот. Элонгация жирных кислот.
пальмитоолеиновая и олеиновая – синтезируются из пальмитиновой и стеариновой кислот.
Наряду с десатурацией жирных кислот (образование двойных связей) в микросомах происходит и их удлинение (элонгация), причем оба эти процесса могут сочетаться и повторяться. Удлинение цепи жирной кислоты происходит путем последовательного присоединения к соответствующему ацил-КоА двууглеродных фрагментов при участии малонил-КоА и НАДФН. Энзиматическая система, катализирующая удлинение жирных кислот, получила название элонгазы. На схеме представлены пути превращения пальмитиновой кислоты в реакциях десатурации и элонгации.

Регуляция синтеза ЖК:
ассоциация/диссоциация комплексов субъединиц фермента Ац-КоА-карбоксилазы. Активатор – цитрат; ингибитор – пальмитоил-КоА.
фосфорилирование/де=//=. Фосфорилированный ф. неактивен(глюкагон и адреналин). Инсулин вызывает дефосфорилирование – становится активной.
индукция синтеза ферментов. Избыт.потребление у/в – ускорение превращения продуктов катаболизма в жиры; голодание или богатая жирами пища приводит к снижении синтеза ферментов и жиров.
Кетонурия и ее причины.
Кетоновые (ацетоновые) тела. В нормальной моче эти соединения встречаются лишь в самых ничтожных количествах (не более 0,01 г в сутки). При выделении больших количеств кетоновых тел качественные пробы становятся положительными. Это явление патологическое и называется кетонурией. Например, при сахарном диабете ежедневно может выделяться до 150 г кетоновых тел.
С мочой никогда не выделяется ацетон без ацетоуксусной кислоты, и наоборот. Обычные нитропруссидные пробы позволяют определить не только присутствие ацетона, но также и ацетоуксусной кислоты; β-оксимас-ляная кислота появляется в моче лишь при сильном увеличении количества кетоновых тел (сахарный диабет и др.).
Кетоновые тела выделяются с мочой не только при сахарном диабете, но и при голодании, исключении углеводов из пищи. Кетонурия наблюдается при заболеваниях, связанных с усиленным расходом углеводов: например, при тиреотоксикозе, кровоизлияниях в подпаутинные пространства, черепно-мозговых травмах. В раннем детском возрасте (продолжительные заболевания пищеварительного тракта (дизентерия, токсикозы) могут вызвать кетонемию и кетонурию в результате голода и истощения. Кетонурия нередко наблюдается при инфекционных заболеваниях: скарлатине, гриппе, туберкулезе, менингите. В этих случаях кетонурия не имеет диагностического значения и является вторичной.
Билет№28
Переваривание и всасывание нуклеопротеинов в ЖКТ. Судьба всосавшихся продуктов.
Переваривание нуклеопротеинов и всасывание продуктов их распада осуществляются в пищеварительном тракте. Под влиянием ферментов желудка, частично соляной кислоты, нуклеопротеины пищи распадаются на полипептиды и нуклеиновые кислоты; первые в кишечнике подвергаются гидролитическому расщеплению до свободных аминокислот. Распад нуклеиновых кислот происходит в тонкой кишке в основном гидролитическим путем под действием ДНК- и РНКазы панкреатического сока. Продуктами реакции при действии РНКазы являются пуриновые и пи-римидиновые мононуклеотиды, смесь ди- и тринуклеотидов и резистентные к действию РНКазы олигонуклеотиды. В результате действия ДНКазы образуются в основном динуклеотиды, олигонуклеотиды и небольшое количество мононуклеотидов. Полный гидролиз нуклеиновых кислот до стадии мононуклеотидов осуществляется, очевидно, другими, менее изученными ферментами (фосфодиэстеразами) слизистой оболочки кишечника.
В отношении дальнейшей судьбы мононуклеотидов существует два предположения. Считают, что мононуклеотиды в кишечнике под действием неспецифических фосфатаз (кислой и щелочной), которые гидролизируют фосфоэфирную связь мононуклеотида («нуклеотидазное» действие), расщепляются с образованием нуклеозидов и фосфорной кислоты и в таком виде всасываются. Согласно второму предположению, мононуклеотиды всасываются, а распад их происходит в клетках слизистой оболочки кишечника. Имеются также доказательства существования в стенке кишечника нуклеотидаз, катализирующих гидролитический распад моно-нуклеотидов. Дальнейший распад образовавшихся нуклеозидов осуществляется внутри клеток слизистой оболочки преимущественно фосфороли-тическим, а не гидролитическим путем.
Всасываются преимущественно нуклеозиды, и в таком виде часть азотистых оснований может быть использована для синтеза нуклеиновых кислот организма. Если происходит дальнейший распад нуклеозидов до свободных пуриновых и пиримидиновых оснований, то гуанин не используется для синтетических целей. Другие основания, как показывают опыты с меченными по азоту аденином и урацилом, в тканях могут включаться в состав нуклеиновых кислот. Однако экспериментальные данные свидетельствуют, что биосинтез азотистых оснований, входящих в состав нуклеиновых кислот органов и тканей, протекает преимущественно, если не целиком, de novo из низкомолекулярных азотистых и безазотистых предшественников.
Биосинтез триацилглицеринов, способы синтеза, последовательность реакций. Роль инсулина, адреналина, глюкагона в регуляции синтеза. Значение процесса.
Известно, что скорость биосинтеза жирных кислот во многом определяется скоростью образования триглицеридов и фосфолипидов, так как свободные жирные кислоты присутствуют в тканях и плазме крови в небольших количествах и в норме не накапливаются.
Синтез триглицеридов происходит из глицерина и жирных кислот (главным образом стеариновой, пальмитиновой и олеиновой). Путь биосинтеза триглицеридов в тканях протекает через образование α-глице-рофосфата (глицерол-3-фосфата) как промежуточного соединения.
В почках, а также в стенке кишечника, где активность фермента глицеролкиназы высока, глицерин фосфорилируется за счет АТФ с образованием глицерол-3-фосфата:
В жировой ткани и мышцах вследствие очень низкой активности глицеролкиназы образование глицерол-3-фосфата в основном связано с процессами гликолиза и гликогенолиза. Известно, что в процессе гли-колитического распада глюкозы образуется дигидроксиацетонфосфат (см. главу 10). Последний в присутствии цитоплазматической глицерол-3-фос-фатдегидрогеназы способен превращаться в глицерол-3-фосфат:
Отмечено, что если содержание глюкозы в жировой ткани понижено (например, при голодании), то образуется лишь незначительное количество глицерол-3-фосфата и освободившиеся в ходе липолиза свободные жирные кислоты не могут быть использованы для ресинтеза триглицеридов, поэтому жирные кислоты покидают жировую ткань. Напротив, активация гликолиза в жировой ткани способствует накоплению в ней триглицеридов, а также входящих в их состав жирных кислот. В печени наблюдаются оба пути образования глицерол-3-фосфата.
Образовавшийся тем или иным путем глицерол-3-фосфат последовательно ацилируется двумя молекулами КоА-производного жирной кислоты (т.е. «активными» формами жирной кислоты – ацил-КоА). В результате образуется фосфатидная кислота (фосфатидат):
Как отмечалось, ацилирование глицерол-3-фосфата протекает последовательно, т.е. в 2 этапа. Сначала глицерол-3-фосфат-ацилтрансфераза катализирует образование лизофосфатидата (1-ацилглицерол-3-фосфата, а затем 1-ацилглицерол-3-фосфат-ацилтрансфераза катализирует образование фосфатидата (1,2-диацилглицерол-3-фосфата) .
Далее фосфатидная кислота гидролизуется фосфатидат-фосфогидро-лазой до 1,2-диглицерида (1,2-диацилглицерола):
Затем 1,2-диглицерид ацилируется третьей молекулой ацил-КоА и превращается в триглицерид (триацилглицерол). Эта реакция катализируется диацилглицерол-ацилтрансферазой:
Синтез триглицеридов (триацилглицеролов) в тканях происходит с учетом двух путей образования глицерол-3-фосфата и возможности синтеза триглицеридов в стенке тонкой кишки из β-моноглицеридов, поступающих из полости кишечника в больших количествах после расщепления пищевых жиров.
Установлено, что большинство ферментов, участвующих в биосинтезе триглицеридов, находятся в эндоплазматическом ретикулуме, и только некоторые, например глицерол-3-фосфат-ацилтрансфераза,– в митохондриях.
Регуляция: Адреналин и норадреналин увеличивают скорость липолиза в жировой ткани; в результате усиливается мобилизация жирных кислот из жировых депо и повышается содержание неэстерифи-цированных жирных кислот в плазме крови. Как отмечалось, тканевые липазы (триглицеридлипаза) существуют в двух взаимопревращающихся формах, одна из которых фосфорилирована и каталитически активна, а другая – нефосфорилирована и неактивна. Адреналин стимулирует через аденилатциклазу синтез цАМФ. В свою очередь цАМФ активирует соответствующую протеинкиназу, которая способствует фосфорилированию липазы, т.е. образованию ее активной формы. Следует заметить, что действие глюкагона на липолитическую систему сходно с действием кате-холаминов.
Гормоны и их классификация. Представления об основных механизмах гормональной регуляции метаболизма.
Г – это химические посредники, регулирующие обмен в-в и развитие органзма.
Биологические признаки:
Дистантность действия
Строгая специфичность, т.е. один Г нельзя заменить другим
Высокая биологическая активность(в крови конц.10-6 – 10-11ммоль/л)
Г оказывают действие путями:
- изменяют проницаемость клеточных мембран
- изменяют скорость ферментативных р-ций – модулируют активность готовых мо-д Б-ферментов, это Г срочной регуляции(сек, мин)
- изменяют скорость синтеза Б-ферментов – Г медленной регуляции.
Классификация:
по химич.природе:
производные АК – адреналин(эпинефрин), норадреналин, тироксин и др.
пептидные(елковые) – АКТГ, ЛТГ, МЦС, инсулин
стероидные – половые, кортикостероиды.
по механизму передачи горм.сигнала в кл-мишени:
стероидные Г и тироксин. Их рецепторы располагаются в цитозоле клеток; они проникают во внутрь и реализуют гормональный эффект по цитозольному механизму.
пептидные и адреналин; рецепторы к этим Г на пов-ти клет.мембран, и Г не проникают вутрь клеток.
в звисимости от посредника:
Г реализуют свой эфф.через циклические нуклеотиды(цАМФ, цГМФ)
с участием ионизированного Са++ и инозитолполифостфатов (ИПФ)
Креатинурия и ее причины.
КРЕАТИНУРИЯ— появление креатина в моче. Моча здорового мужчины почти не содержит креатина, у женщин и детей — незначительная физиологическая креатинурия; у женщин она увеличивается при беременности, лактации. Избыточное содержание в пище мяса может привести к экзогенной креатинурией.
Основной причиной патологической креатинурии являются поражения мышц: миозиты, мышечная дистрофия, тяжелая миастения, тонические и клонические судороги.
Кроме того, креатинурия отмечается при ряде эндокринных заболеваний — диабете, гипертиреозе, акромегалии, а также при ацидозе, алкалозе, авитаминозах Е и С.
В норме образующийся в мышцах креатин ангидрируется н выделяется с мочой в виде креатинпна (около 2 г в сутки). Т. к. возможность организма ангидрировать креатин ограничена, возникновение креатинурии может иметь место как при повышенном распаде белка, так и при нарушении нормального превращения креатина в креатинин.
Определение креатина в моче производят путем превращения его в креатинин при помощи гидролиза 1 н. раствором соляной кислоты. Определяют количество креатинина в моче реакцией Яффе до и после гидролиза. Вычитанием первой величины из второй узнают содержание в моче креатина.
Билет№29
Пути распада пуриновых и пиримидиновых нуклеотидов в тканях. Конечные продукты. Нарушения обмена нуклеотидов. Биохимические основы подагры.
Распад пуриновых: Пиримидиновых:
конечные продукты – СО2, аммоний
и бета-аланин(бета-аминоизомасляная к-та):
-трансаминирование(в альфа-ала)
-исп-ся в мышцах на образование дипептидов –
ансерина и карнозина
- под действием м/флоры кишечника
включ.в пантотеновую к-ту – на образ.КоА
Превращение и всасывание липидов в желудочно-кишечном тракте.
Сут. потребность 180г. Должны быть представлены в разных кол-вах насыщенные, полиненасыщенные и мононасыщенные. 30% по калорийности от всей пищи. ХС в сут.должно быть менее 300мг.
рНопт. липазы жел.сока =5,5-7,5, а у взрос. 1,5, поэтому расщепление в желудке не происходит(в кишечнике!)
У груд.детей рНжел.сока 5,0. Л.расщепляются лингвальной липазой(рНопт 4,5)и желудочной липазой.
Основные фазы превращения:
Эмульгирование – образование из большиз липидных капель мелкодисперсной эмульсии(0,5мкм). Э.способствует: *натриевые сили желчных кислот, *СО2 – образ-ся при разложении бикарбонатов под действием соляной к., поступающей с пищевым комком из желудка, *образующиеся по мере расщеп.липидов мыла(соли жир.к-т и натрия R-COONa)
Осн.роль в эмульгировании у желчи – смеси натриевых солей желчных к-т, ФЛ и ХС желчных к-т в соотн. 12,5 : 2,5 : 1.
рН желчи больше 7,0.
Желчные к-ты – конечный продукт обмена ХС – производные холановой к-ты:
Желчные кислоты:
- первичные 3,7,12-триоксихолановая = холевая; 3,7-диоксиходановая = хенодезоксихолевая(образ.из печени из ХС)
- вторичные 3,12-диоксихолановая = дезоксихолевая; 3-оксихолановая=литохолевая(образ.в кишечнике под действием м/флоры)
**Конъюгирование желчных кислот: 1)с глицином – при у/в питании; 2)с таурином(образ.из цистеина)-при Б питании
роль: - эмульгируют жиры; -активируют липазу, -способсвуют всасыванию продуктов гидролиза жиров в кишечнике.
В сутки из ХС образ. 1г желчи в гепатоцитах→по печеночным пузырным протокам→концентр.в желч.пузыре.Опорожнение происходит постоянно, но усил.под действием поступившей пищи и ХЦкинина.
После выполенния своих ф.85%желчных кислот всас.в подвздошной кишке и вновь поступ.в состав секретируемой желчи – энтерогепатальный круг.
Общий пул желч.к-т 2,8-3,5г. Совершают 6-8 оборотов в сут.
Недостаток желч.к-т: желчекам.болезнь, стеаторрея.
Липолитическая фаза.
Осн.продукты – бета-моноацилглицерин и жир.к-ты.
Эфиры ХС(ферм.холестерол-эстераза панкреатич.и кишеч.соков) →жир+жир.к-ты.
Фосфатидилхолин (ф.панкреатич.фосфолипаза А2) →лизофосфолипид(панкреатич.лизофосфолипаза) →жир.к-та(из альфа)+глицерофосфохолин→всас.в кровь или выводятся.
Мицеллярная фаза. Транспортировка продуктов расщепления липидов от места образования к всас.пов-ти кишечника. Глицерин и жир.к-ты с Сменьше 10 всас-ся хорошо. Жир.к-ты с числом С-атомов больше 10, ХС, нерасщепленные ФЛ всас.с помощью мицелл. Они в сотни раз меньше эмульсий. В центрк гидрофобное ядро из жир.к-ты, моноглицерида и ХС. Снаружи гидрофильн.оболочка из ФЛ и жнлч.к-т.
мукозная фаза. Процессы ресинтеза в клетках эпителия кишечника – бета-моноглицеридный синтез ТАГ: поскольку жир.к-ты поступили извне(с пищей), из них образ.экзогенные:
R-COOH + HSKoA + АТФ (1)→ RCOSKoA + АМФ + ФФн
бета-МАГ + R1COSKoA(2) → ДАГ (3)→ ТАГ
Триглицеридсинтетаза: (1)ацилКоАсинтетаза, (2)моноглицеридацилтрансфераза, (3)диглицеридацилтрансфераза.
5) Транспортная фаза. Трансп.форма липидов – ЛП. Л, образованные в энтероцитах тонкогг киш. – экзогенные ТАГ, а также ФЛ и всосавшийся ХС – окружаются белком и формир. ХМ – очень крупные частицы, не способные проникать в кров. капилляры, поэтому диффундируют в димфатич.с-му кишечника→грудной лимфатич.проток. Легкие ограничивают поступление ХМ в арт.кровь(липопектическая ф.легких). ХМ не облад. атерогенностью: из-за больших размеров не проникают в капилляры. После приема пищи наблюдается гипергликемия(алиментарная) за счет ХМ. Х гидролизуются ЛП-липазой(крепится к стенкам сосудов гепарином)
Гормоны щитовидной и паращитовидной желез. Химическое строение и участие в обменных процессах.
тироксин. Фолликулярная часть щит.ж. 3,5,3’,5’-тетрайодтиронин
Регулирует скорость основного обмена, рост и дифференцировку тканей, обмен белков, у/ви липидов, водно-солевой обмен, деят-ть ЦНС, ЖКТ…
гипофункция: в детском возрасте – кретинизм, остановка роста, изменения кожи, волос. мышц, наруш.психики.
Гпотиреоидный отек(микседема). Слизистый отек, ожирение, выпад.волос и зубов, психич.расстройства.
Эндемический зоб(недостаток иода)
Гиперфункция: гипертиреоз – диффуз.токсич.зоб/Базедова болезнь: тахикардия, пучеглазие+зоб, общее истощение+психич.расстройства.
паратгормон. Паращитовид.ж. Белковая природа: 84 АК-остатка;состоит из 1 п/п цепи. Физиологическое влияние паратГ на клетки почек кост.ткани через систему аденилатциклаза-цАМФ. Регуляция конц.Са++ и связанных с ним анионов фосфотной к-ты в крови. В почках уменьш.реабс.Са++
Гипоф: тетанические судороги
Гиперф: вымывание солей Са в виде цитратов и фосфатов из костной ткани→деструкция мин.и орг. компонентов костей.
3) кальцитонин. С-клетки щит.ж. Пептидная природа. Выделяется в ответ на повышение Са++ в крови. Пост. конц. Са в крови → уменьш.резорбции кост.тк., гипокальциемия, гипофосфатемия – антагонист паратгормона.
Протеинурия и ее причины.
почечная
функциональная – временное проходящее нарушение почечного фильтра. Физиогогическая – у новорожд., обусловлена увелич. прониц-тью поч.фильтра; к 4-10 дню проходит. Ортостатическая(циклич.) – у детей 6-12 лет и подростков(приходящие изменения гемодинамики почки, например, из-за аномалии осанки). Инсулярная – у детей раннего возр.(повыш.раздражимость почесного фильтра)
органическая. Гломерулярная – поврежд.почеч.фильтра(селективная – в моче Б с массой менее 100кДа, и неселективная – в моче все Б крови). Тубулярная – поврежд.канальцев и наруш.работоспособности Б.
внепочечная
преренальная/перегрузочная – увелич. конц.общ.Б или появление парапротеинов в плазме: легкие цепи иммуноглоб(Б Бенс-Джонса), Нв(ППВ=1,25г/л), миоглобин(раюдомиолиз)
постренальная – попадание в мочу Б из мочевывод.путей при воспал.или опухолях.
Билет№30
Биосинтез ДНК. ДНК-полимеразы. Повреждения и репарация ДНК. Наследственные заболевания, связанные с нарушением репарации ДНК.
Буферные системы крови. Роль буферных систем в поддержании гомеостаза pH. Кислотно-основное состояние. Понятие об ацидозе и алкалозе.
В организме кислотообразование преобл.над образованием соед-й основного характера.
Источники Н+ в организме:
1.летучая кислота Н2СО3, в сутки образ.10-20тыс.ммоль СО2 при окислении белков, Ж, У.
2.нелетучие кислоты в сут.образ. 70 ммоль:
- фосфорная при расщепл.орг.фосфатов(нуклеотидов, ФЛ, фосфопротеидов)
-серная, соляная при окислении Б
3.орг.к-ты:молочная, кетоновые тела, ПВК и др.
рН удерживается на слабощелочном уровне благодаря участию буф.с-м и физиологическому контролю(выделит.ф.почек и дыхат.ф.легких)
Уравнение Гендерсона-Хессельбаха: pH = pKa + lg [акц.протонов]/[донор протонов].
( Соль) ( кислота)
Любая буф.с-ма состоит из сопряженной кислотно-основной пары: донор+акцептор протонов.
Буферная емкость: зависит от абсолютных концентраций компонентов буфера.
Бикарбонатная.
10%буф.емкости крови.
![]()
При нормальном значении рН крови (7,4) концентрация ионов бикарбоната НСО3 в плазме крови превышает концентрацию СО2примерно в 20 раз. Бикарбонатная буферная система функционирует как эффективный регулятор в области рН 7,4.
Механизм действия данной системы заключается в том, что при выделении в кровь относительно больших количеств кислых продуктов водородные ионы Н+ взаимодействуют с ионами бикарбоната НСО3–, что приводит к образованию слабодиссоциирующей угольной кислоты Н2СО3. Последующее снижение концентрации Н2СО3 достигается в результате ускоренного выделения СО2 через легкие в результате их гипервентиляции (напомним, что концентрация Н2СО3 в плазме крови определяется давлением СО2 в альвеолярной газовой смеси).
Если в крови увеличивается количество оснований, то они, взаимодействуя со слабой угольной кислотой, образуют ионы бикарбоната иводу. При этом не происходит сколько-нибудь заметных сдвигов в величине рН. Кроме того, для сохранения нормального соотношения между компонентами буферной системы в этом случае подключаются физиологические механизмы регуляции кислотно-основногоравновесия: происходит задержка в плазме крови некоторого количества СО2 в результате гиповентиляции легких.
NaHCO3 + H+ → Na+ + H2CO3
Реабс.в почках ↓карбоангидраза
СО2 + Н2О
↓увеличение вентиляции легких
Фосфатная представляет собой сопряженную кислотно-основную пару, состоящую из иона Н2РО4– (донорпротонов) и иона НРО42– (акцептор протонов):
![]() Фосфатная буферная
система составляет
всего лишь 1% от буферной емкости крови.
Во
внеклеточной жидкости,
в том числе в крови,
соотношение [НРО42–]:
[Н2РО4–]
составляет 4:1.
Буферное
действие фосфатной системы основано
на возможности связывания
водородных ионов ионами НРО42– с
образованием Н2РО4– (Н+ +
+ НРО42– —>
Н2РО4–),
а также ионов ОН– с ионами Н2РО4– (ОН– +
+ Н2 Р
О4– —>
HPO42–+
H2O).
Буферная пара (Н2РО4––НРО42–)
способна оказывать влияние при изменениях
рН в интервале от 6,1 до 7,7 и может
обеспечивать определенную буферную
емкость внутриклеточной жидкости,
величина рН которой в пределах 6,9–7,4.
В крови максимальная
емкость фосфатного буфера проявляется
вблизи значения рН 7,2.
Фосфатная буферная
система составляет
всего лишь 1% от буферной емкости крови.
Во
внеклеточной жидкости,
в том числе в крови,
соотношение [НРО42–]:
[Н2РО4–]
составляет 4:1.
Буферное
действие фосфатной системы основано
на возможности связывания
водородных ионов ионами НРО42– с
образованием Н2РО4– (Н+ +
+ НРО42– —>
Н2РО4–),
а также ионов ОН– с ионами Н2РО4– (ОН– +
+ Н2 Р
О4– —>
HPO42–+
H2O).
Буферная пара (Н2РО4––НРО42–)
способна оказывать влияние при изменениях
рН в интервале от 6,1 до 7,7 и может
обеспечивать определенную буферную
емкость внутриклеточной жидкости,
величина рН которой в пределах 6,9–7,4.
В крови максимальная
емкость фосфатного буфера проявляется
вблизи значения рН 7,2.
1 и 2 – выводящие.
Белковая имеет меньшее значение для поддержания КОР в плазме крови, чем другие буферные системы. Белки образуют буферную систему благодаря наличию кислотно-основных групп в молекуле белков: белок–Н+ (кислота, донорпротонов) и белок (сопряженное основание, акцептор протонов). Белковая буферная система плазмы крови эффективна в области значений рН 7,2–7,4.
Гемоглобиновая буферная система – самая мощная буферная система крови. Она в 9 раз мощнее бикарбонатного буфера; на ее долю приходится 75% от всей буферной емкости крови. состоит из неионизированного гемоглобина ННb (слабая органическая кислота, донорпротонов) и калиевой соли гемоглобина КНb (сопряженное основание, акцептор протонов). Точно так же может быть рассмотрена оксигемоглобиновая буферная система. Система гемоглобина и система оксигемоглобина являются вза-имопревращающимися системами и существуют как единое целое.
Механизм действия:
- в тканях: Н2О + СО2(карбоангидраза) —>Н2СО3—>Н+ + НСО3-(диффундирует в плазму крови)
КНвО2 —>КНв + 4О2
КНв + 2Н+ —> ННв + 2К+ (К-гемоглобин неитрализует ионы Н+)
- в легких: ННв + 4О2 —>2Н+ + НвО2
2Н+ + НвО2 + 2К+ + 2НСО3- —>КНвО2 + 2Н2СО3(карбоангидраза) —>Н2О + 2СО2
рН и концентрация СО2 влияют на освобождение и связывание О2 немоглобином – эфф.Бора.
Повышение концентрации протонов, СО2, способствует освобождению О2, а повышение концентрации О2 стимулирует высвобождение СО2 и протонов.
Гормоны надпочечников. Глюкокортикоиды и минералокортикоиды. Химическое строение и участие в обменных процессах.
Кортизол/гидрокортизон – гормон пучковой зоны коры надпочечников. Глюкокортикоид/кортикостероид.
Влияние на обмен у/в, белков, жиров и нукл.к-т. Стимулирует образование глю в печени, усил.глюконеогенез, синтез глк в печени, и уменьш.потребление глю периферическими тканями. Липолиз в конечностях и липогенез в теле.
Альдостерон - клубочковая зона коры. Минералокортикоид.
Гл.образом регулир.обмен натрия, калия, хлора, воды. Способств. Удержанию натрия и хлора в организме и выведению с мочой ионов калия.
Нарушения:
Первичная недостаточность – б.Аддисона(поражение коры). Снижение массы тела, слабость, снижение аппетита, тошнота, снижение АД, «бронзовая болезнь»
Вторичная – отсутствует гиперпигментация кожи, -//-.
Врожденная гиперплазия надпочечников. Нарушение синтеза кортизола – усиление роста тела, раннее половое созревание у мальчиков и муж.пол.признаки у девочек.
Гипекортицизм(при опуходях гипофиза.). Гипергликемия, уменьш.толерантности к глю, уменьш.мыш.массы, истончение кожи, остеопороз.
Гематурия и гемоглобинурия, их причины.
Гематурия:
Ренальная – поражение эпителия канальцев или интерстиция почки
Постренальная – травматическое повреждение мочевыводящих путей(например, отхождение камней)
Выделяют следующие формы гемоглобинурий: 1. Ночная гемоглобинурия Маркиафава. 2. Токсические гемоглобинурии. 3. Маршевая гемоглобинурия (редкое заболевание).
После долгих переходов (маршей) может наблюдаться типичная гемоглобинурия. Всегда имеется выраженный поясничный лордоз. Других патологических явлений нет. Гемоглобинурия через несколько часов прекращается. Особого клинического значения эта форма не имеет.
4. Холодовая гемоглобинурия: при пароксизмальной холодовой гемоглобинурии, в большинстве случаев после длительного воздействия холода (купание в холодной воде, реже только холодный наружный воздух), наблюдается гемоглобинурия.
Билет№31
Биосинтез РНК. Процессинг матричной и транспортной РНК. Обратная транскрипция, биологическая роль.
Транскрипция – синтез РНК на матрице ДНК.
Последовательность дезоксирибонуклеотидов в гене ДНК переписывается в виде комплементарной последовательности РНК.
Кодирующая – с которой пишется.
Транскриптон – участок ДНК, на котором происходит синтез РНК: *стр-рные гены, кодирующие белки, * промотор, * терминатор.
Синтез РНК катализирует ДНК-зависимая РНК-полимераза. Холофермент РНК-полимеразы состоит из 5 субъединиц: α, α’, β, β’, δ. РНК-полимераза без δ – корфермент(ядро). Субъединица δ нужна для узнавания промотора и связывания с ним.
Этапы транскрипции:
Связывание РНК-полимеразы с ДНК
Холофермент РНК-пол.двигается по ДНК, пока не найдет промотор, образуя с ним сначала закрытый промоторный комплекс. Чтобы информация стала открытой, он должен стать открытым. Расплетается с участием δ. Связь РНК-пол.с промотором становится прочнее.
Инициация
Состоит в образовании первых звеньев в цепи РНК. Субстраты и источники Е – макроэргические рибонуклеозидтрифосфаты(АТФ, ГТФ, УТФ, ЦТФ)
РНК-полимеразы не треб.затравки(в отл.от ДНК-пол). Первый нуклеотид вход.в состав цепи в виде трифосфата;следующий присоед.к его 3’-группе.
Синтез РНК идет комплементарно и антипараллельно кодирующей цепи РНК. Когда образована цепь из 3-9 нуклеотидов, дельта-субъединица отщепляется.
Стадию элог\нгации и терминации катализирует корфермент.
На стадии элонгации расплетено 18 пар нуклеотидов;12 из них образуют гибридную спираль с растущей ДНК.
РНК-полимераза двигается по ДНК; впереди нее водород.связи разрываются, сзади замыкаются.
РНК-пол.не проверяет правильность синтеза;ошибки чаще, чем при ДНК.
Терминация
Участок ДНК перед терминатором богат цитозиновыми и гуаниловыми нуклеотидами.
Шпилька останавливает ДНК-полимеразу;транскрипция заканчивается. РНК-полимераза и синтезированная РНК отделяются от ДНК. Терминации способствуют белки-факторы терминации.
Транскрипция идет консервативно – ДНК-матрица полностью сохраняется; РНК синтезируется заново.
РНК-пол.у эукариот:
РНК-пол.1 – синт.в ядрышке рибосомальную РНК
РНК-пол.2 – синт. Информационную РНК
РНК-пол.3 – транспортные и др.РНК
Синтезированная РНК(первичный транскрипт) не может выполнять свою биологич.ф., т.к. является незрелой. Для ее созревания нужен процессинг
У прокариот не происходит. Транляция начинается еще до завершения транскрипции
У прокариот включает модификацию 5’-конца(кэпирование) и 3’-конца(полиаденилирование)
Обратная транскрипция.
Синтез ДНК на матрице РНК.
Характерна для ретровирусов. Имеют фермент ревертазу(обратная транскриптаза). Используют в качестве матрицы как ДНК, так и РНК.
В клетке хозяина ревертаза вируса синтезирует цепь ДНК, комплементарную матрице РНК. Затем расщепляет всю вирусную РНК. На основе ДНК происходит синтез второй ДНК. Далее двойная ДНК встраивается в кл-хозяина.
Гемоглобин, строение и свойства. Возрастные особенности. Понятие об аномальных гемоглобинах.

В крови 130-160г/л(у муж.132-164, жен.115-145г/л), в эритр.300-400г/л(в ед.СИ)
Нв – хромопротеин(не энзимный гемопротеин). Небелковая часть – гемм, и белковая – глобин.
Имеет 4 уровня стр-ры:
первичная – послед-ть АК в п/п цепях, попарно одикановых. Так. НвА(взрос) имеет 2альфа и 2бета-цепи. Альфа-цепь имеент 141 АК, бета – 146, и вся мол-ла =574АК.
Спиральзация п/п цепей. Мол-ла Нв спирализована на 80%, и представляет правозакрученную спираль. Альфа-цепь имеет 7 спиральных участков., бета – 8. Спир.участки обознач.буквами лат.алф. – А,В,С…Н. Они чередуются с неспирализованными, обозначаемыми двумя буквами спиралей, между кот.они находятся(АВ,ВС)
П/п цепи образ.множнство связей. В отдельных местах спиральные участки делают изгибы, повороты, образуя третич.стр-ру:
упаковка спирали в простр-ве. При этом гемм оказ-ся окутанным в слое белка. При упаковке цепей глобина неполярные АК оказ-ся внутри(окружая гем) придавая высокую устойчивость железу гемма(защищ.от окисл-я)
Каждая п/п цепь связана с гемом: гемм+глобин = субъед-ца/протомер
Связь осущ.через железо гемма и атом азота гистидина(в п/п цепи). п/п цепи образ.для гемма карманы, окруженные неполярными АК и ужерживают. Железо гемма имеет координационное число 6: 4 связи направлены к 4 атомам азота пиррольных колец гемма, 5я – к атому азота гистидина в п/п цепи, 6я – на связь с О2(в дезоксиНв она свободна).
ассоциация протомеров, ориентированных в пространстве определенным образом относительно друг друга. Вся мол-ла Нв имеет 4 гема и 4 глобина(п/п цепи)
Между субъед-цами возникает множ-во контактов – α1-α2, β1-β2, α 1-β1, α 2-β2…
Эти контакты удерживают м-лу Нв в виде димера и тетрамера.
Субъед-цы ассоциированы ак, что в центре образ.впадина – центр.полость, в к-рой фиксируются орг.фосфаты.
Свойства:
*транспорт О2 к тканям (через железо гемма-простетич.гр)
*транспорт СО2 из тканей (через своб.амынные гр.АК-белк.часть)
Высокая р-римость СО2(в 20 раз больше чем у О2)создает благоприят.усл.для транспорта, к-рый осущ.своб.аминн.гр.глобина – карбаминоНв.
*обеспеч.поддержание постоянства рН, входя в состав гемоглобиновой буф.с-мы
*выполн.антитоксич.ф. – связ.цианиды – цианметНв, монооксид углерода – карбоксиНв.
Гетерогенность Нв.
Онтогенетическая/созревания.
Смена типов зависит от:
-особенностей снабжения тканей О2. у эмбриона источник – интерстиц.жидкость, у плода – плацента, у новорожденного – легкие.
- смены органов кроветворения
1. эмбриональные Нв. Их синтез идет с 2х недель эмбр.развития;ощутимые кол-ва опр-ся в 8 нед.
(Gower-1(эпсилон2,эпсилон2), Gower-2(эпсилон4), Portland-1(гамма2,епсилон2))
Синтез прекращ.к 14 нед.(образование плаценты)
фетальные. HbF(альфа2,гамма2)-гетерогенен(3-5компонентов)
свойства: лучше р-рим, чем НвА;высокая скорость окисления железа гемма – легко происх.образоване метНв. Устойчив к денатурации щелочью. Обладают более высоким сродством к О2. имеет специфич.спектр поглощения. Высокая скорость при электрофорезе. Синтезируется с 14нед.внутриутр.разв.
возрастные изменения: н/р 80%,3мес 40%,6мес 23%,9мес 10%, 1год 4-5%, 3года 1,5%, больше3лет 1%.
2)Нв взрослого типа.
Синтез НвА начинается у эмбр.на 8-11нед., идет медленно и у н/р сост.20%от общего. 1мес 43%, 5мес 75%.
При комплексировании Нв с у/в. Конмлекс аминогр.с альд.гр. глю образ.А1с, образуется альдиминная связь, к-рая при длительной гликемии превращ.в стойкую кетоаминную с образованием гликированных Нв.
Функции почек. Транспорт веществ в процессе секреции и реабсорбции. Реабсорбция глюкозы, аминокислот, профильтровавшихся белков. Пороговые и беспороговые вещества.
Экскреция и мочеобразование - выведение конечных продуктов обмена, чужерод.в-в, изб.Н2О, мин. и орг. солей.
гомеостатическая – регуляция стабильности ионного состава и осм.давления в вод.секторах, ОЦК, КОС и АД.
Инкреторная – синтез биологич.акт.соед., регулир.ф.самих почек и оказ.влияние на метаболизм в целом:
-эритропоэтин/ингибитор эритропоэза
-вит.Д3-кальцитриол
-ренин-протеолитический фермент
-тромбопластин – сверт.кр.
- урокиназа – запуск сист.фибринолиза
- колликреин-кининовая сист.
-простагландины(горм.широкого спектра действия – влияют на поч.кровоток, водно-солевой обмен, тонус сосудов)
4) Метаболическая. особенности:
- образ. аммиака
- синтез бикарбоатов
- синтез глю(глюконеогенез) – особое знач.в условиях голодания и при ацидозе(10%в норме образ.в почках)
- катаболизм профильтрованных пептидов и низкомол-лярных Б(удаление Б., утративших нативную стр-ру, удаление изб.ферментов(амилаза), предотвращение потери АК)
азотемия, гипертония, анемия, остеодистрофия, наруш.водно-сол.обмена, отеки, сдвиги рН.
Функц.активность почек просих.в нефроне:ультрафильтрация, реабс-я, секреция, синтез новых соед.
Ультрафильтрация.
Осущ.пассивно а клубочках путем просачивания жид.ч.крови через капилляры с образованием первич.мочи(ультрафильтрата). В сут. ок.180л. По химич. сост.он аналогичен плазме, за искл.Б. В норме филтруется 2-4г Б за сутки.(М меньше 50-60кДа)
причины ограничения фильтрации Б:
небольшой диаметр поч.фильтра
одноим. «-»заряд почеч.фильтра(увелич.содерж.гепарпнсульфатов) и Б плазмы кр., обр-е электростатич.батьера.
В ультрафильтрате могут присутствовать амилаза, лизоцим, альбумины, альфа1, альфа2-глою., легкие цепи Ig.
Колич.хар-кой фильтрации явл. скорость жен. – 110мл/мин
муж. – 12мл/мин.
измерение скорости проводят по клиренсу ряда неабсорбируемых.соед. – инсулин, креатинин.
С=U/Р*V, С-клиренс, И-конц. в-ва в моче, Р-конц.в-ва в плазме, - минутный диурез.
С креатинина=145мл/мин.
Механизмы транспорта в-в в ходе реабсорбции и секреции.
Трансцеллюлярный(через кл)
парацеллюлярный(через м/Кл контакты)натрий, вода.
Движущими силами переноса в-в явл.концентр.и электрохимич.градиенты. перенос может очущ.путем пассивног транспорта.
Акт.трансп.:натрий-калиевая АТФаза →вторич.
↓
акт.трансп.с участием Б-переносчиков и ионами:натрий+-симпорт: глю, АК, орг.А-, Cl-, K+, Pn, HCO3-, антипорт: Н+
↓
эндоцитоз и трансцитоз, хар-ный для Б(ецептор-зависимый механизм)
Пассивный транспорт:дифф.СО2, Н2О, хлора и др.
Реабсорбция.
Обратное всас.в кровь; в проксим. канальцах всас. 65%воды, натрия и хлоридов, 100%глю, АК, вит.К. В дист.канальцах всас.осмотич.своб.вода, электролиты, кальций, фосфаты(регуляция с помощью Г – альдостерон, АДГ, паратГ, кальцитонин, кальцитриол)
беспороговые в-ва: конц.в моче пропорциональна накоплению в крови
Слабоабсорбируемы в-ва: сочевая к-та, мочевина.
Неабсорбируемые: креатинин, инсулин.
Пороговые – хорошо реабс-ся и имеют порого почеч.выведения.
ППВ – наим.конц.в-ва в крови, при к-рой оно появл.в моче. Глю, АК, б/р, креатин, фосфаты, бикарбонаты, Б, Нв. ППВ для альбуминов=60-70г/л.
Секреция.
(в прокс.канальцах) Актив.транспор. осуществляемый в разных направлнеиях.
1. из крови в просвет канальцев(ксенобиотики, орг.в-ва и к-ты, калий+)
2. из эпителия канальца в просвет(иммиак+, Н+)
3. из эпителия в кровь(НСО3-)
В рез-те образ.вторич.моча. Значит.отличие от плазмы по химич.составу: орг. в-ва(35-45г/сут), азотистые(мочевина. креатинин, креатин), парные соед-я(живот.индикан, гиппуровая к-та, фенол и крезол – глюкурон.к-ты), ЩУК, лимонная. укс.к-ты, БАВ(вит.и Г), мин.в-ва(15-25г/сут: кат+:натрий, калий, аммоний, кальций, магний, Анионы-: схлор, сульфат, фосфат)
Фенилкетонурия, алкаптонурия. Причины их возникновения.
ФКУ. фенилпировиноградная олигофрения. Развивается как ре-т потери способности организма синтезировать фенилаланин-4-монооксигеназу, катализирующую превращение фен и тир. Резкое замедление умственного развития ребенка, экскреция с мочой большого кол-ва фенил ПВК(до 1-2г/сут), и фенилацетилглутамина(2-3г/сут), накопление фен в тканях. Его кол-во в крови при норме 15мг/л – до 600мг/л, в СМЖ до 80 при норме 1,5. Предотвращение: исключение фен с пищей с самого рождения.
фен→ фен-гидроксилаза × Тир×катехолаины
↓ ×тиреоид.Г
фенил-ПВК → фенилацетат ×меланины
→ фениллактат
Алкаптонурия. Экскреуия с мочой больших кол-в гомогентизиновой к-ты(до 0,5г/сут), окисление которой О2 воздуха придает моче темную окр. Охроноз, отложение пигмента в тканях и потемнение носа, ушей, склеры. Врожд.дефект связан с отсутствием печени и почках оксидазы гомогентизиновой к-ты.
Билет№32
Биосинтез пуриновых нуклеотидов. Роль фолиевой кислоты. Синтез дезоксирибонуклеотидов, роль системы тиоредоксина. Синтез нуклеотидтрифосфатов.
Роисхождение атомов пуринового кольца:
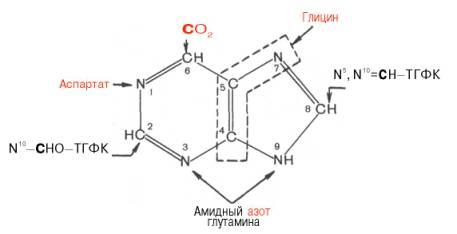
Из схемы видно, что 4-й и 5-й атомы углерода и 7-й атом азота в ядре имеют своим источником глицин. Два атома азота (N-3 и N-9) происходят из амидной группы глутамина, один атом азота (N-1) – из азота аспара-гиновой кислоты; углеродный атом (С-2) происходит из углерода N10-фор-мил-ТГФК, атом углерода в 8-м положении – из N5,N10-метенил-ТГФК и, наконец, углерод С-6 имеет своим источником СО2.
АМФ и ГМФ образуются из ИМФ, причем в синтезе обоих моно-нуклеотидов участвуют по два фермента, различных по своему механизму действия. Образование ГМФ из ИМФ катализируют ИМФ-дегидрогеназа и ГМФ-синтетаза, а образование АМФ из того же предшественника катализируется последовательным действием аденилосукцинатсинтетазы и аденилосукцинат-лиазы. Механизм двухэтапного синтеза АМФ и ГМФ можно представить в виде химических реакций.
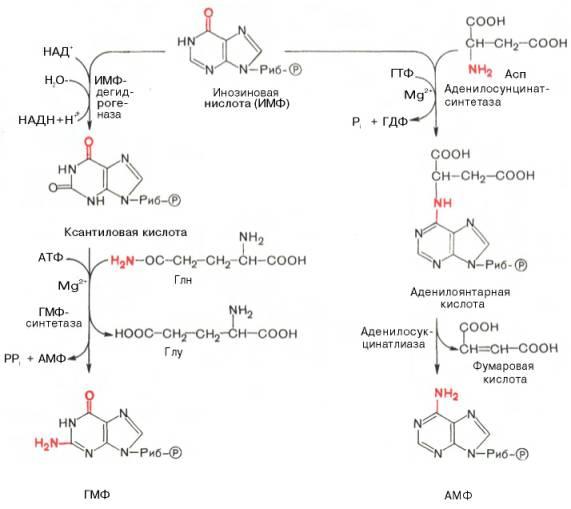
В ферментативном синтезе АМФ из ИМФ специфическое участие принимает аспарагиновая кислота, являющаяся донором NH2-группы, и ГТФ в качестве источника энергии; промежуточным продуктом реакции является аденилоянтарная кислота. Биосинтез ГМФ, напротив, начинается с де-гидрогеназной реакции ИМФ с образованием ксантозиловой кислоты; в аминировании последней используется только амидный азот глутамина.
Превращение АМФ и ГМФ в соответствующие нуклеозидди- и нуклео-зидтрифосфаты также протекает в 2 стадии при участии специфических нуклеозидмонофосфат- и нуклеозиддифосфаткиназ :
ГМФ + АТФ <=> ГДФ + АДФ; ГДФ + АТФ <=> ГТФ + АДФ.
Фолиевая кислота. Вит.Вс

Ее активная форма – ТГФК нужна на этапе синтеза ИМФ.
Синтез дезоксирибонуклеотидов из нуклеотидтрифосфатов
В самом начале процесса происходит потеря рибонуклеозидтрифосфатами одной фосфатной группы и образуются АДФ, ГДФ, ЦДФ, УДФ.
Во второй реакции фермент рибонуклеозид-редуктаза восстанавливает АДФ, ГДФ, ЦДФ, УДФ до дезоксирибонуклеозиддифосфатов dАДФ, dГДФ, dЦДФ, dУДФ. Донором водорода для восстановления рибозы является белок тиоредоксин, его SH-группы окисляются кислородом рибозы и образуется вода. Последующее восстановление тиоредоксина в рабочее состояние обеспечивается за счет НАДФН.
После образования dАДФ, dГДФ, dЦДФ фосфорилируются, а dУДФ используется для синтеза тимидилового нуклеотида.
Депонирование и мобилизация жиров в жировой ткани.
Жиры, как и гликоген, являются формами депонирования энергетического материала. Причем жиры — наиболее долговременные и более эффективные источники энергии . При голодании запасы жира у человека истощаются за 5—7 недель, тогда как гликоген полностью расходуется примерно за сутки. Если поступление жира превышает потребности организма в энергии , то жир депонируется в адипоцитах — специализированных клетках жировой ткани. Кроме того, если количество поступающих углеводов больше, чем надо для депонирования в виде гликогена, то часть глюкозы также превращается в жиры . Таким образом, жиры в жировой ткани накапливаются в результате трех процессов:
1.поступают из хиломикронов, которые приносят экзогенные жиры из кишечника
2.поступают из ЛОНП, которые транспортируют эндогенные жиры, синтезированные в печени из глюкозы
3.образуются из глюкозы в самих клетках жировой ткани.
В первом и во втором случае жиры в составе липопротеинов гидролизуются ЛП—липазой и в клетку поступают жирные кислоты, которые затем используются для синтеза ТАГ. Перед включением в ТАГ жирные кислоты сначала активируются путем образования тиоэфиров кофермента А, а затем взаимодействуют с глицеролфосфатом. Так как глицерол не может быть фосфорилирован в адипоцитах (в этих клетках отсутствует глицеролкиназа), то глицеролфосфат образуется при восстановлении диоксиацетонфосфата.
Следовательно, синтез ТАГ может протекать только в присутствии глюкозы, из которой в процессе гликолиза образуется диаксиацетонфосфат. Инсулин стимулирует синтез ТАГ, потому что в его присутствии повышается проницаемость мембран клеток жировой ткани для глюкозы.
Мобилизацию (липолиз) депонированных ТАГ катализирует тканевая липаза. В результате жиры распадаются на глицерин и свободные жирные кислоты.
Адреналин и глюкагон активируют внутриклеточную липазу. Действие этих гормонов опосредовано аденилатциклазным каскадом реакций, начиная с активации аденилатциклазы и заканчивая фосфорилированием липазы, которая при этом переходит в активную форму и расщепляет эфирные связи в ТАГ. Глицерол как растворимое в плазме вещество транспортируется в печень, где используется в реакциях глюконеогенеза. Жирные кислоты транспортируются кровью в виде комплексов с сывороточными альбуминами в разные органы и ткани, где включаются в процесс окисления.
Ферменты сыворотки крови. Классификация. Диагностическое значение их определения.
1)Плазмоспецифические(секреторные)
2)Экскреторные
3)Индикаторные(клеточные)
Активность ферментов определяется для правильной постановки топического диагноза; для проведения дифференциальной диагностики; для оценки эффективности лечения; для определения прогноза у пациента.
Гиперферментемия:
увеличение проницаемости клет.мембран
деструкция клет.мембран
активация синтеза белков-ферментов
нарушение выведения белков-ферментов с желчью или мочой.
Гипоферментемия – снижение активности в пл.крови:
активный захват ферментов клетками РЭС
нарушение синтеза б-ферментов вследствие алиментарных причин (белковое голодвние), недостаточеого поступления витаминов, микроэл-тов(Fe, Cu, Mg, Mn..), выполняющих роль коферментов.
Плазмаспецифические.
Синтезируются в печени; биологич.роль выполняют в плазме крови, поэтому диагностич.значение имеет только понижение активности данных ф.
ЛХАТ лецитин-холестерин-ацил-трансфераза
Осущ.р.этерификации ХС в стр-ре альфа-липопротеинов.
ЛП-липаза(ф-р просветления)
Уч.в гидролизе триглицеридов в стр-ре ХМ и ЛПОНП(пре-бета-ЛП)
Псевдохолинэстераза
Участ.в расщеп.ацетилхолина, бутирилхолина. Диагностич.значение имеет при б.Боткина и при отравлении фосфоорганич.соединениями(активность резко снижается)
Б. Лизоцим – фактор неспецифической резистентности организма
Церулоплазмин(медь-содержащая оксидаза)
Экскреторные ферменты.
Синтезируются в печени, поджел., слюн.железах. физиологич.роль выполняют в ЖКТ. Диагностич.значение имеет только повыш.активности: поступ.в кровь, где и обнаруживаются.
Альфа-амилаза. Осущ.гидролиз крахмала и глк (через стадии образования декстринов до мальтозы)
Повыш.: патологии поджел.ж., слюн.железы(паротит), заболев.легких.
Щелочная фосфатаза(фосфомоноэстераза первого типа)
Ф.образуется в печени;выходит в ЖКТ, где осущ.гидролиз моноэфиров.
Увелич.: закупорка желчевыв.протоков(ж/к болезнь), патологии костной системы
Лейцин-аминопептидаза. Ф.поджед.железы, гидролизующий короткоцепочечные пептиды. Увелич.: пат.подж.ж.(наруш.процесс выхода в ЖКТ)
Индикаторные ферменты.
Диагностич. Значение имеет только повыш.активности. роль выполняют непосредственно в клетке. Вследствие нарушения проницаемости клет.мембран она обнаруж-ся в плазме крови.
Цитоплазматические:
- АлАТ
- КФК и ее изоферменты (креатин-киназа, ММ, МВ, ВВ)
- лактат-ДГ и ее изоферменты
- альдолаза
Митохондриальные:
- АсАТ
- орнитин-карбамоил-трансфераза
- цитохром-оксидаза
- глутамат-ДГ
Лизосомы:
- протеолитические белки-катепсины h, b, l-классы
- кислые фосфатазы
- виды нуклеаз(ДНКаза, РНКаза)
Роль воды в организме. Содержание и распределение воды в тканях. Возрастные особенности. Регуляция водного обмена.
Вода организма образует 2 водных пространства: внутриклеточное(2/3общей воды) и внеклеточное(1/3). Небольшое кол-во воды входит в жидкости полостей тела: боюшной, плевральной и т.п., а также в цереброспинальную, анутриглазную, внутрисуставную жид.. Внеклеточное пр-во включает: 1) внутрисосудистый вод.сектор, т.е. плазму крови, объем к-рой сост.8%общей воды, и 2) интерстициальный вод.сектор, содержащий ¼ всей воды(15%массы тела) и являющийся наиб.подвижным при избытке или недостатке воды в теле.
25%-интерстициальная жидкость
8%-плазма (внутрисосудистый фактор)
67%-внутриклеточная жидкость.
Вода является основным веществом, из которого состоит организм человека.
Содержание воды в теле зависит от возраста, пола, массы тела. У здорового взрослого мужчины массой 70 кг общее содержание воды в организме составляет около 60% массы тела, т.е. 42 л. У женщин общее количество воды в организме приближается к 50% массы тела, т.е. меньше, чем у мужчин, что обусловлено большим содержанием бедной водой жировой ткани и меньшим – мышечной. У новорожденного ребенка содержание воды в теле достигает 80% массы тела и затем с возрастом постепенно уменьшается вплоть до старости. Это – одно из проявлений старческой инволюции, зависящее от изменения свойств коллоидных систем (снижение cпособности молекул белков связывать воду) и от возрастного уменьшения клеточной массы, главным образом мышечной ткани. Общее содержание воды зависит также от массы тела: у тучных оно меньше, чем у лиц с нормальной массой тела, у худых – больше. Это связано с тем, что в жировой ткани воды значительно меньше, чем в тощих тканях (не содержащих жира).
Вода выполняет важные функции в организме:
- Она является обязательной составной частью всех клеток и тканей.
- Выступает в роли универсального растворителя органических и неорганических веществ.
- В водной среде протекает большинство химических реакций, т.е. процессов обмена веществ,
лежащих в основе жизнедеятельности организма.
- Она участвует в транспорте субстратов, необходимых для клеточного метаболизма, и выведении из организма вредных продуктов обмена веществ.
- Вода определяет физико-химическое состояние коллоидных систем, в частности дисперсность белков, что обусловливает их функциональные особенности.
Регуляция
Осмотическая регуляция.
Поступление воды в организм обеспечивается изменением питьевого поведения, которое зависит от чувства жажды. Центр жажды составляют нейроны ряда областей головного мозга: образований, окружающих 3-й желудочек, преоптической зоны и гипоталамуса. Их особенностью является наличие больших вакуолей, заполненных жидкостью с определенной осмоляльностью. Таким образом, в формировании жажды принимают участие различные системы гипоталамических нейронов, интегрирующие сенсорные влияния, обусловленные видом и вкусом воды с внутренними сигналами, идущими от осморецепторов, локализованных в перижелудочковой, преоптической зонах и в гипоталамусе. Система сравнения, расположенная около нейронов СОЯ, может контролировать их активность и таким образом оказывать влияния на высвобождение АДГ
Волюмическая регуляция, важнейший механизм сохранения вводно-электролитного баланса, также участвует в формировании жажды. Волюморецепторы каротидного синуса и дуги аорты и барорецепторы
предсердий, устьев полых вен и легочной вены реагируют соответственно на изменения объема и давления циркулирующей крови.
В реализации осмо- и волюморефлексов - основных инструментов регуляции осмотического гомеостаза участвуют гуморальные механизмы.
Гуморальная регуляция вводно-электролитного обмена осуществляется гормонами и местно образующимися физиологически активными веществами, в частности простагландинами Е. Основными гормонами, регулирующими осмотический гомеостаз, являются вазопрессин (АДГ), гормоны РААС
(ангиотензин-2 и альдостерон), ПНУФ (атриопептин).
Аргинин – вазопрессин (ВП), или антидиуретический гормон (АДГ), представляет собой нонапептид, образующийся нейроэндокринными нейронами супраоптического (СОЯ), паравентрикулярного (ПВЯ) и ряда добавочных ядер гипоталамуса.
Активация ВП-нейронов возникает при увеличении осмоляльности плазмы крови на 1-2 % и при гиповолемии за счет импульсов от осмо- и волюморецепторов.
Основной эффект ВП можно свести к эуосмии - поддержанию осмотического давления крови и тем самым межклеточной жидкости, что создает условия для сохранения стабильного объема клеток в организме, определяющего их эффективное функционирование, и установлению соответствия объема
циркулирующей крови емкости сосудистого русла.
Главным органом-мишенью для гормонов – регуляторов водно-электролитного обмена являются почки. Основное значение ВП связано с регуляцией конечного этапа реабсорбции воды в почечных канальцах.
Билет№33
Биосинтез пиримидиновых нуклеотидов. Особенности синтеза тимидиловых нуклеотидов, тимидилатсинтетаза, роль тетрагидрофолиевой кислоты (ТГФК). Нарушения синтеза пиримидиновых нуклеотидов.
В клетках животных и микроорганизмов промежуточными продуктами синтеза не являются свободные пиримидиновые основания, и остаток рибозы присоединяется к уже сформировавшемуся пиримидиновому кольцу.
Последовательность химических реакций синтеза пиримидиновых нук-леотидов, в частности УМФ, можно представить в следующем виде:
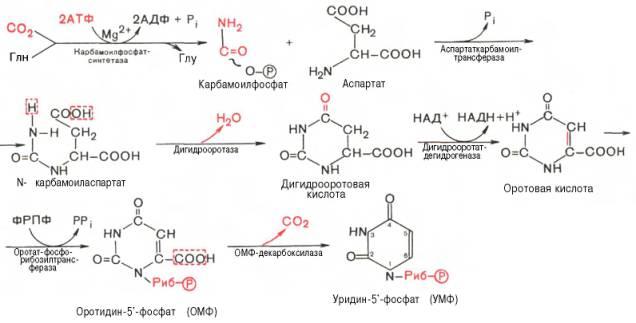
Биосинтез тимидиловых нуклеотидов. Тимидиловые нуклеотиды входят в состав ДНК, содержащей дезоксирибозу. Поэтому сначала рассмотрим механизмы синтеза дезоксирибонуклеотидов. При помощи метода меченых атомов было показано, что этот синтез начинается не со свободной дезоксирибозы, а путем прямого восстановления рибонуклеотидов у 2'-го атома углерода. При инкубации меченых предшественников (рибонуклео-тидов) в бесклеточной системе бактерий метку обнаружили в составе дезоксирибонуклеотидов. По данным П. Рейхарда, у Е. coli все 4 рибо-нуклеозиддифосфата восстанавливаются в соответствующие дезоксиана-логи: dАДФ, dГДФ, dЦДФ, dУДФ – при участии сложной ферментной системы, состоящей по меньшей мере из четырех разных ферментов.
Химический смысл превращения рибонуклеотидов в дезоксирибо-нуклеотиды сводится к элементарному акту – восстановлению рибозы в 2-дезоксирибозу, требующему наличия двух атомов водорода. Непосредственным источником последних оказался восстановленный термостабильный белок тиоредоксин, содержащий две свободные SH-группы на 108 аминокислотных остатков. Тиоредоксин легко окисляется, превращаясь в дисульфидную S-S-форму. Для его восстановления в системе имеется специфический ФАД-содержащий фермент тиоредоксинредуктаза (мол. масса 68000), требующая наличия восстановленного НАДФН. Обозначив условно рибонуклеозиддифосфат РДФ, образование дезоксирибонуклео-тидов можно представить следующим образом:
Обе стадии могут быть представлены в виде схемы:
Для синтеза тимидиловых нуклеотидов, помимо дезоксирибозы, требуется также метилированное производное урацила – тимин. Оказалось, что в клетках имеется особый фермент тимидилатсинтаза, катализирующая метилирование не свободного урацила, а dУМФ; реакция протекает по уравнению:
Донором метильной группы в тимидилатсинтазной реакции является N5,N10-метилен-ТГФК, которая одновременно отдает и водородный протон, поэтому одним из конечных продуктов реакции является не тетра-гидро-, а дигидрофолиевая кислота (ДГФК). Последняя вновь восстанавливается до ТГФК под действием НАДФН-зависимой дигидрофолат-редуктазы. Из образовавшегося ТМФ путем фосфотрансферазных реакций образуются dТДФ и dTТФ.
Синтез всех остальных дезоксирибонуклеозид-5'-трифосфатов, непосредственно участвующих в синтезе ДНК, также осуществляется путем фосфорилирования дезоксирибонуклеозид-5'-дифосфатов в присутствии АТФ:
АТФ + dАДФ –> АДФ + dATФ; АТФ + dЦДФ –> АДФ + dЦТФ;
АТФ + dГДФ –> АДФ + dГТФ; АТФ + dТДФ –> АДФ + dТТФ.
Механизмы защиты от активных форм кислорода. Ферментные и неферментные звенья антиоксидантной защиты.
Молекулярный кислород сам по себе обычно не вступает в неконтролируемые химические реакции внутри организма, для его активации нужны ферментативные процессы – главные ферменты метаболизма кислорода у млекопитающих: оксидазы и оксигеназы. Но в каталитических центрах этих ферментов кислород испытывает превращения до конечных соединений, не выделяясь в среду и не подвергая опасности органические макромолекулы клетки, повреждающими же агентами являются активные формы кислорода (АФК), образующиеся в ряде физико-химических процессов в организме.
Во всех аэробных клетках в процессе присоединения одного электрона к молекуле кислорода образуются супероксидный анион-радикал - О2- и его протонированная форма – гидроперекисный радикал - НО2*; оба они порождают ряд других активных форм кислорода. Образование этих АФК наиболее существенно вблизи цепей переноса электронов – дыхательная цепь, микросомы и, в растительных клетках, хлоропласты.
Гидроперекиси липидов являеются весьма активными соединениями и обладают высокой биологической агрессивностью. Для протекания цепного окисления липидов в биологических мембранах совершенно необходимы переходные металлы, в частности, ионы железа.
Простым и доступным методом определения продуктов перекисного окисления липидов является реакция с тиобарбитуровой кислотой (Marcuse, Johnston, 1973). Главным механизмом защиты организма от данных форм АФК является фермент супероксиддисмутаза (СОД), активность ее обычно достаточна, чтобы инактивировать их в месте образования, не допуская диффузии в среде макромолекул ткани.
Дисмутация супероксидных анион-радикалов под действием СОД в биологических тканях ведет к образованию перекиси водорода, способной легко проникать через мембраны клеток. Перекись водорода обнаруживается при фагоцитозе, при работе митохондрий и микросом. В присутствии ионов переходных металлов (например Fe2+) перекись водорода может давать высоко активный гидроксильный радикал (* ОН). Этому процессу препятствуют главные высоко активные ферменты антиоксидантной защиты организма: каталаза и глутатион-пероксидаза.
Высокой реакционной способностью обладает гидроксильный радикал (* ОН), образующийся из перекиси водорода в присутствии ионов переходных металлов. Высокая реакционная способность определяет преимущественно местное воздействие этой форм АФК. Прямое повреждение ДНК при этом характеризуется разрывом цепи (180); с другими биомолекулами * ОН образует вторичные свободные радикалы, в том числе перексиные соединения липидов.
Главные типы повреждений биомолекул * ОН: отрыв атома водорода (таким образом повреждается лецитин – главный компонент биологических мембран, а также сахара в составе нуклеозидов ДНК); присоединение к молекулам по двойным связям (взаимодействие с пуринами и пиримидинами ДНК и РНК, в том числе с образованием вторичных радикалов); перенос электронов также является патогенным механизмом действия * ОН.
ЗАЩИТА ОРГАНИЗМА ОТ АФК
СУПЕРОКСИДДИСМУТАЗА (СОД). Супероксиддисмутаза является важнейшим элементом антиоксидантной защиты организма. Это фермент из двух субъединиц с общей молекулярной массой 32 кДа, содержащий по одному атому меди и цинка (существует также марганец-содержащая СОД, обнаруженная в печени крысы и человека; в бактериальных клетках обнаружена железо-содержащая СОД). Фермент ускоряет распад О2- на 4 порядка.
Источником О2- обычно является система феназинметасульфат + НАД*Н или ксантин + ксантиноксидаза. За единицу активности СОД принято ее количество, тормозящее в 2 раза восстановление цитохрома с при реакции ксантиноксидазы (0,003 ед/мл), ксантина (2 мкмоль/мл) и цитохрома с - 0,6 мкмоль/мл
КАТАЛАЗА. Каталаза расщепляет перекись водорода, до которой дисмутирует супероксидный радикал, до молекул воды и молекулярного кислорода. В клетках каталаза в основном сосредоточена в пероксисомах, в которых содержатся и ферменты, продуцирующие перекись водорода, необходимую в ходе ряда процессов жизнедеятельности организма, в частности, в процессах неспецифической иммунной защиты.
ПЕРОКСИДАЗА. Пероксидаза, в особенности глутатион-пероксидаза, широко распространена в клетках животных и растений. Глутатион-пероксидаза состоит из 4 субъединиц в каждой из которых содержится по молекуле селена. В клектах этот фермент располагается в цитозоле и матриксе митохондрий.
Активность глутатион-пероксидазы зависит от содержания глутатиона клетки, что, в свою очередь, определяется активностью глутатионредуктазы и концентрацией НАДФ*Н, который образуется в пентозофосфатном метаболическом цикле. Лимитирующими органами по активности каталазы, являются легкие, мышцы, глаза.
ДРУГИЕ ФОРМЫ ЗАЩИТЫ ОТ АФК
В защите от АФК в организме участвуют и многие другие молекулы и ферментные системы.
Аскорбиновая кислота инактивирует свободные радикалы, образуя неактивный радикал (семидегидроаскорбат), она же является кофактором пероксидазы, (фермент аскорбат-пероксидаза).
Глутатион, присутствуя в клетках в высоких концентрациях, также является акцептором гидроксильного иона и синглетного кислорода, кроме того, он же является кофактором глутатион-пероксидазы и глутатион-редуктазы. Мочевая кислота присутствует в крови в достаточных количествах, чтобы эффективно акцептировать синглетный кислород и гидроксильный радикал. Аналогичными эффектами обладают этанол, маннит, глюкоза и некоторые другие органические молекулы.
Пантотеновая кислота. Химическая природа, распространения, участие в обменных процессах.
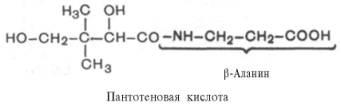 2,4-диокси-3,3-диметилмасляная
кислота
2,4-диокси-3,3-диметилмасляная
кислота
Витамин В5/антидерматитный. Суточная потребность – 3-5мг; источник – печень, дрожжи, желток, зеленые части растений. Активная форма – коэнзим А(ацетилированная форма кофермента А)
Биол. роль пантотеновой кислоты обусловлена ее участием в биосинтезе кофермента А (КоА, KoASH; ф-ла II)-мол.м. 767,54; бесцв. кристаллы; хорошо раств. в воде; 260 нм (рН 2); 14,6·103. КоА-акцептор и переносчик разл. кислотных остатков (см. Коферменты). Реакционноспособной частью молекулы КоА является концевая сульфгидрильная группа. Ацильные группы присоединяются к коферменту при помощи тиоэфирной связи. Своб. энергия гидролиза ацил-КоА имеет большую отрицат. величину (напр., для гидролиза ацетил-КоА она составляет 31,4 кДж/моль). Таким образом кофермент переносит ацильные группы, подобно тому как АТФ переносит активир. фосфорильные группы. Для КоА характерно тиолдисульфидное взаимод. (RS—SR + + R'SHRS—SR' + RSH), а также регуляторные воздействия на ряд ферментативных р-ций (особенно на регулирующих соотношение КоА/ацил-КоА).
Гиповитаминоз: дерматиты, поражения слизистых оболочек, дистрофии желез внутр.секреции(надпочечники) и НС(невриты, параличи), изменения в сердце и почках, депигментация волос, прекращение роста, потеря аппетита.
Гомеостатическая функция почек. Участие почек в регуляции КОС. Процессы ацидо- и аммониогенеза. Титруемая кислотность мочи. Аммонийные соли.
Роль почек в осмо- и волюморегуляции. Почки являются основным органом осморегуляции. Они обеспечивают выделение избытка воды из организма в виде гипотонической мочи при увеличенном содержании воды (гипергидратация) или экономят воду и экскретируют мочу, гипертоническую по отношению к крови, при обезвоживании организма (дегидратация).
После питья воды или при ее избытке в организме снижается концентрация растворенных осмотически активных веществ в крови и падает ее осмоляльность. Это уменьшает активность центральных осморецепторов, расположенных в области супраоптического ядра гипоталамуса, а также периферических осморецепторов, имеющихся в печени, почке и других органах, что приводит к снижению секреции АДГ нейрогипофизом и увеличению выделения воды почкой.
При обезвоживании организма или введении в сосудистое русло гипертонического раствора NaCl увеличивается концентрация осмотически активных веществ в плазме крови, возбуждаются осморецепторы, усиливается секреция АДГ, возрастает всасывание воды в канальцах, уменьшается мочеотделение и выделяется осмотически концентрированная моча.
Помимо осмо- и натриорецепторов, уровень секреции АДГ определяет активность волюморецепторов, воспринимающих изменение объема внутрисосудистой и внеклеточной жидкости.
Роль почек в регуляции ионного состава крови. Почки являются эффекторным органом системы ионного гомеостаза. В организме существуют системы регуляции баланса каждого из ионов. Для некоторых ионов уже описаны специфические рецепторы, например натриорецепторы. Рефлекторная регуляция транспорта ионов в почечных канальцах осуществляется как периферическими, так и центральными нервными механизмами.
Роль почек в регуляции кислотно-основного состояния.
1)поддержание нормальной концентрации бикарбонатов крови: полная реабсорбция бикарбоната; образование доп.кол-в бикарбоната.
2)избирательная экскреция: оснований(в форме бикарбонатов и двузамен.фосфатов), кислот(в форме титруемых к-т и аммонийных солей)
Образование аммонийных солей.
Кровь просвет канальца
NaCl-←NaCl
Na+ ←Na+ ↓
HCO3←NCO3-←H2CO3→H+→ H+→NH4
Глутамин(1)→глутамат → 2*NH3→ NH3→
↓(2)
альфа-КГ ↓
NH4Cl
экскреция 30-50ммоль
(1)глутаминаза, (2)глутамат-ДГ.
Аммиак в просвете - буфер.осн.для связывания ионов Н+ с образованием аммонийного иона – аммониогенез. рН мочи в норме = 6,0 – 6,5.
Ацидогенез
Ацидогенез. Роль почек в сохранении КОС заключается в выведении из кислой крови Н + и из основной — НСО3. В клетках почечных канальцев под воздействием карбоангидразы происходит реакция
СО2 + Н2О -> Н2СО3
Н2СО3 диссоциирует на Н+ и НСО3В канальцевую мочу поступает бикарбонат натрия, который диссоциирует на Na и НСО3. Н+ из клеток почечных канальцев поступают в канальцевую мочу и, соединясь с НСО3, образуют Н2СО3, из которой диссоциируют Н2О и СО2. В свою очередь, натрий из канальцевой мочи поступает в канальцевые клетки и, соединяясь с НСО3, в виде бикарбоната натрия поступает в венозную кровь капилляров канальцев. СО2, образовавшийся в канальцевой моче, поступает в канальцевые клетки, где также под влиянием карбоангидразы и реакции С02 + Н20 образуется угольная кислота Н2СО3, которая диссоциирует на Н+ и НСО3
В канальцевой моче содержится соль фосфата натрия (Na2HPO4), которая диссоциирует на Na+ и НРО4. Na из канальцевой клетки перемещается в канальцевую мочу, где образуется NaH2PO4 — кислая соль фосфата натрия, которая выводится с мочой. Na+ из канальцевой мочи перемещается в канальцевые клетки, где соединяется с НСО3 и в виде NaHCO3 поступает в венозную кровь капилляров канальцев.
Титруемую кислотность мочи определяют титрованием ее 0,1 н. NaOH. Титрование продолжают до тех пор, пока рН мочи не достигнет величины рН плазмы крови (7,4). Конец титрования определяют выравниванием цвета мочи с исходным цветом буфера (в состав буфера входит моно- и дифосфат). Израсходованное на титрование количество щелочи делят на 10, умножают на суточное количество мочи. Таким образом, определяют количество титруемых кислот, выделенных за сутки в мл 0,1 н. раствора NaOH.
Билет№34
Распад хромопротеинов в тканях. Фазы превращений билирубина. Исследование желчных пигментов с диагностической целью.
У взрослого человека 700-900г гемоглобина; ежесут.распад.0,01часть, т.е 8-9г. Нв превращ.в желчные пигменты.
Осн. Поставщик – эритроциты, которые живут 120дней;при их разруш. Гемогл. →в кровь.
В крови Нв встреч.с белком гаптоглобином(Нр):
Это гемопротеин, транспорт-ся с альфа-2-глобул.фракцией, имеет 2 центра связывания с Нв и в комплексе с Нв не вывод.с мочой
Нр состоит из 4 п/п цепей – 2альфа и 2бета – и имеют 3 нег.типа: Нр1-1, Нр1-2, Нр2-2
Нр строго индивид.для каждого человека. Его исп.для установления отцовства, материнства и т.п.
Нв-Нр комплекс→в кл.РЭС-гистиоциты, кл.Купфера, кл.кр.к.мозга.
Стадии превращения Нв в желчные пигменты:
1)В клетках РЭС
Под действ.гем-оксигеназ.системы(НАДФ, вит.С, тетрагидрофолиевая к-та) превращается в вердоглобин.
Нв+Нр→комплекс
→О2, НАДФН2 ↓Р450, ТГФК, вит.С, гемоксигеназа, →гаптоглобин
НАДФ, Н2О←
Вердоглобин
Fe3+← ↓→глобин→СО
Биливердин – 1й желч.пигмент, зеленый, не токсичный, хорошо раств.в воде.
Биливердин- ↓восст-е
редуктаза,
НАДФН2
Билирубин – пигмент красно-корич.цвета, токсичен, нераств.в воде;обнаруж.в крови – непрямой, несвязанный, неконъюгированный. В крови абсорбирован на альбуминах. В комплексе с последним теряет токсичность, становится хорошо растворим в воде, но с мочой не выводится.
Непрямой несвяз.б/р – для его обнаруж. Необх.осадить Б спиртом или кофеином, затем провести диазореакцию. Он образуется к летках РЭС.
2) Превращение б/р в гепатоцитах.
Идет в 3 фазы:
1. Б\р подходит к гепатоциту, отсоед-ся от альбумина и при участии y и z-протеинов(спец.Б лигандины)поступают в цитозоль гепатоцита, затем в ЭПС
2. В ЭПС гепатоцита: б/р конъюгтрует с УДФГК(акт.фотма глюкурон.к-ты) б/р+УДФГК→БМГ(моно-)→БДГ(ди-)
3. 75% БДГ поступает в желч.пузырь против градиента конц., т.е. это Е-завис.транспорт. Из ж.п. →тон.кишечниу. До 25% БДГ пост.в кровь.
Обнаруж. В крови б/р прямой конъюгированный связанный. Прямой – т.к. он сразу реагирует с диазор-вом Эрлихаю В абс.величинах уровень б/рне превыш. 5 мкмоль/л. Конъюг.-с глюкурон.к-той, возник.хим.свзь;глюкур.к-та присоед.через остатки пропионовой к-ты. Происходит уменьш. Токсичности путем увелич-я р-римости.
3) Превращ.б/р в ЖКТ
БДГ
В тон. ↓→глюк.к-та
кишке
мезобилирубин
восст-е↓редуктазы норм.м/флоры
мезобилиноген(уробилиноген) – его часть всас.через вена порта, поступ.в печень, а там разруш.до пирролов, монодипирролов, а они снова возвращ.в кишечник или вывод. с мочой.
↓
Cтеркобилиноген →с содержимым кишечника и окраш.его; при участии воздуха окисл.в стеркобилин.
(нормальный пигмент кала и мочи)
↓
Через с-му геморроидальных вен всас. →в большой круг→вывод.с мочой
При патологии в печени нарушается превращение мезобилиногена(уробилиногена)в пирролы, и он поступает в кровь, оказывая токсическое действие на ЦНС и др., а затем выводится с мочой. Т.о., это патологический комп.мочи.
Клинический интерес представляет:
С мочой выводится повыш.содерж.уробилиногена→недостаточность функции печени(печеночная или гемолитическая желтуха – потеря печенью способности извлекать пигмент из крови воротной вены)
Исчезновение стеркобилиногена из мочи при наличии б/р и биливердина→полное прекращение поступления желчи в кишечник(при желчекаменной болезни, рак.поражениях поджел.железы)
Биосинтез холестерина, последовательность реакций до образования мевалоновой кислоты, представление о дальнейших этапах. Транспорт холестерина. Холестерин и атеросклероз.
Происходит в цитозоле клеток. Можно выделить 3 этапа:
1)Образование мевалоната.
2 мол-лы Ац-КоА конденсир-ся тиолазой →ацетоАц-КоА. Гидроксиметилглутарил-КоА-синтаза присоед-ет третий ацетильный остаток с обр-ем ГМГ-КоА(3-гидрокси-3-метилглутарил-КоА). Далее под действием ГМГ-КоА-редуктазы идет восстановление ГМГ-КоА до мевалоната с использованием 2 мол. НАДФН.
2)Образование сквалена.
Мевалонат превращ. В пятиуглеродную изопреноидную стр-ру, содержащую пирофосфат – изопентенилпирофосфат, 2 мол-лы которого конденсируются с обр. геранилпирофосфата. Присоединение еще одной изопрен.единицы→фарнезилпирофосфат(15 С-ат), 2 мол-лы которого конденсир-ся с обр. сквалена(30 С-атомов, имеет линейную стр-ру)
3)Образование холестерола
Сквален(ф.циклаза)→ланостерол(4 конденсированных цикла и 30 С-ат.) →…20р-ций.. →холестерол(27 С-ат)