

Класс Двудольные. Classis Ditfotyledones
(грен, di, dis — два, дважды; kotyledon — ёпадина, полость)
Класс двудольных включает около 170 000 видов, что составляет около 75% всех цветковых. Это разнообразные травы, кустарники, лианы и деревья. Общим для всех является наличие двух семядолей, четырех- или пятичленный цветок (пять чашелистиков, пять лепестков, пять тычинок), листья в подавляющем большинстве случаев с сетчатым жилкованием.
Листья двудольных простые или сложные. Простые листья состоят из одной пластинки, а сложные из нескольких, расположенных перисто или веерно вдоль общей оси (рис. 70). Простые
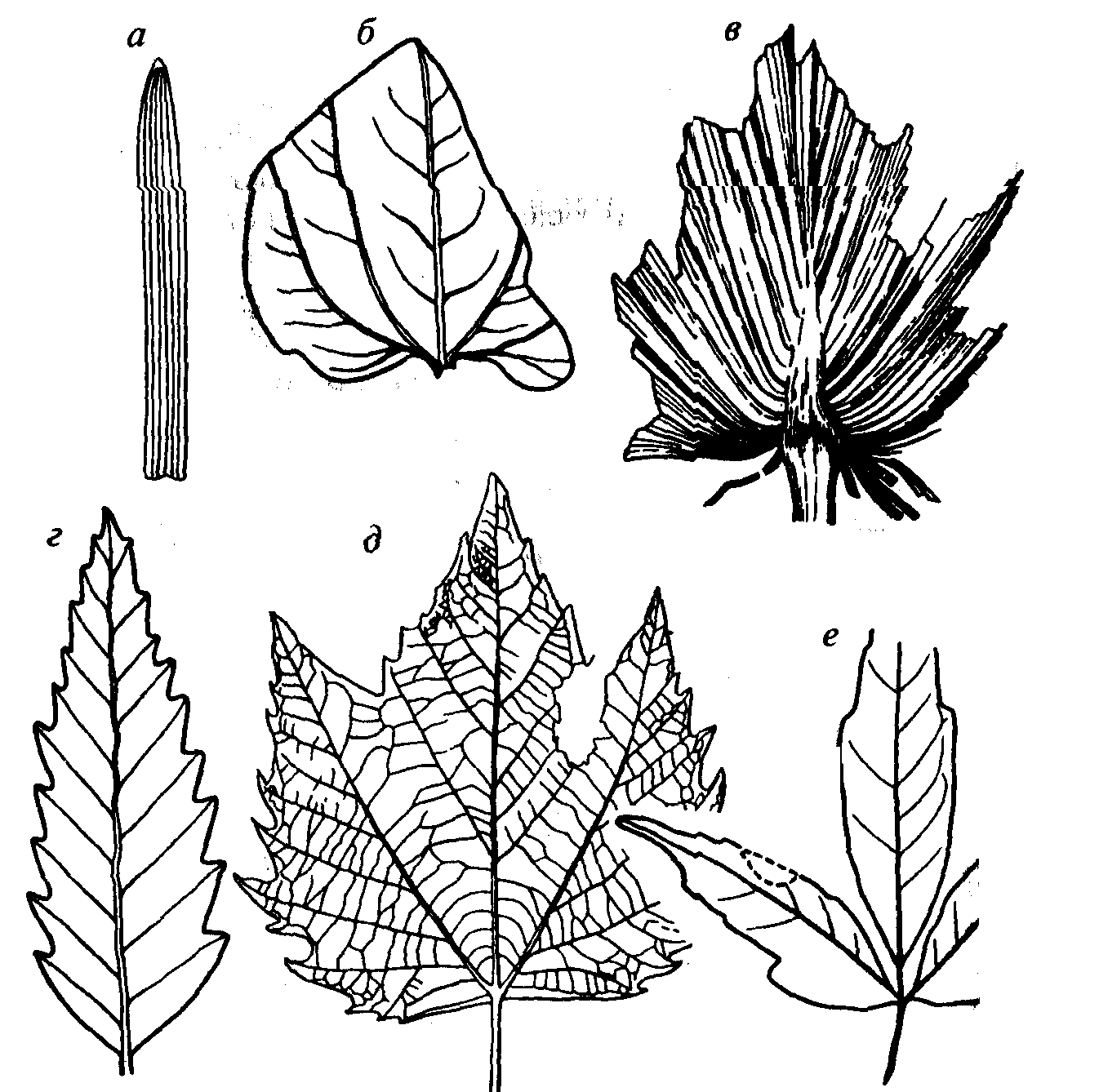
Рис. 70. Отдел Magnoliophyta (K.-Q)
а-в — листья однодольных: а — злаковые, лист с параллельным жилкованием, б— Smilax (K2-Q). лист с дугонервным жилкованием; в — Sabal (P). лист с веерным жилкованием: г-е — листья двудольных: г — Quercus (K2-Q), д — Platanus (K-Q), e - Rhus (K2-Q)
листья встречаются как у травянистых растений, так и у деревьев. Сложные листья характерны для деревьев и в меньшей степени для трав. Жилкование сетчатое с хорошо выраженными анастомозами; в основном перистое, пальчатое, веерное с жилками нескольких порядков, реже — параллельное, дуговидное. Для классификации листьев, особенно ископаемых, чрезвычайно важентип жилкования.
Мел — современность.
Класс Однодольные. Classis Monocotyledones
(грен, monos — один; /cotyledon — впадина, полость)
Однодольные насчитывают 65 000 видов, т.е. по численности резко уступают двудольным. Они представлены преимущественно травянистыми растениями (злаки, водные и болотные формы), редко кустарниковыми и древовидными растениями (пальмы). Общим для всех является наличие у зародыша одной семядоли, цветок, как правило, трехчленный. Форма листьев вытянутая, кинжалоподобная, лентовидная (лилии, злаки, водные растения), реже стреловидная, эллиптическая (ландыш), перистая либо веерная (пальмы). Жилкование преимущественно параллельное, дуговидное, веерное, реже сетчатое, обычно со слабо выражен- ными анастомозами (см. рис. 70).
Мел — современность.
Геологическая история и породообразующая роль растений
Единичные, несколько сомнительные находки растительного происхождения известны начиная с раннего рифея (1700 млн лет); с венда (670 млн лет) можно уверенно говорить о существовании водорослей, хотя достоверные находки зеленых и красных водорослей известны с кембрия.
Родственные связи и история развития полцарства низших растений дискуссионны. Освоение растениями суши стало возможно благодаря тому, что предки наземных растений достигли определенного морфофизиологического уровня, позволяющего существовать в наземных условиях, сначала в переходной зоне — литорали и на побережьях, а затем и на обширных пространствах континентов. Развитие высших растений шло по пути становления тканей и органов. К сожалению, по ранним этапам освоения суши имеется скудный и неоднозначный геологический материал. Скорее всего предки большинства современных наземных растений возникли в силуре. В докембрии и раннем палеозое влагоемкие пространства суши, возможно, уже были освоены моховидными.
Долгое время полагали, что предками первых высших наземных растений — риниофитов — были бурые водоросли, в настоящее время предпочтение отдают зеленым водорослям.
Риниофиты (S-D) были представлены травянистыми, преимущественно полуводными, реже наземными растениями. Нижняя часть стебля находилась в воде, а верхняя — возвышалась над водой. Риниофиты появились в первой половине силура и заселили побережья окружающих морей и океанов, лагуны, а позднее долины рек и болотистые низины (рис. 71). Их расцвет приурочен к раннему-среднему девону. Риниофиты, возможно, принимали участие в формировании девонских горючих сланцев и углей.
Плауновидные (S2?, D-Q) в девоне, а возможно, в позднем силуре, возникли от риниофитов. Первые плауновидные — небольшие растения, а в карбоне и перми — в основном высокие деревья. Многие древесные плауновидные имели воздухоносные полости, что свидетельствует о произрастании в манграх — влажных тропических лесах, расположенных вдоль побережья морских бассейнов. Наибольший расцвет плауновидных приурочен к карбону, когда они являлись основными углеобразователями.

Рис. 71. Родословное древо высших растений
Папоротниковидные (D2-Q) произошли от риниофитов в среднем девоне; в карбоне и юре они дважды испытали расцвет. Помимо травянистых форм они представлены древесными растениями и лианами, имеющими настоящие листья, корни и довольно сложные спорангии. Папоротниковидные наряду с голосеменными были основными углеобразователями юрского времени.
Хвощевидные (D3-Q) произошли от риниофитов в позднем девоне. Они были представлены травами, деревьями и лианами, имеющими членистый стебель, настоящие листья и корни. Наибольшего разнообразия они достигли в карбоне, особенно в зарослях типа мангровых. В карбоне хвощевидные вместе с плауновидными и папоротниковидными были основными углеобразователями.
В конце девона от папоротниковидных возникли пинофиты, или голосеменные (D3-Q). Они представлены кустарниками, деревьями, лианами, крайне редко травянистыми формами. Начиная с перми число и разнообразие голосеменных резко увеличиваются, достигая максимума в юре. Они основные компоненты углей юрского периода совместно с папоротниковидными.
В раннем мелу появляются цветковые растения (K-Q), вероятно, возникшие от беннеттитовых, но не исключено, что от каких-то других голосеменных. Сначала появились двудольные, а потом и однодольные. Цветковые представлены всеми жизненными формами: травами, кустарниками, полукустарниками, деревьями, лианами. Травяной покров с палеогена и поныне почти целиком составляют цветковые; это в значительной мере относится и к современным лесам, где древесным и кустарниковым цветковым принадлежит ведущая роль, хотя местами они могут уступать хвойным, а также мхам и папоротникам. В неогене цветковые растения участвовали в образовании промышленных угольных пластов (Сахалин).
Чрезвычайно велика роль ископаемых растений для расчленения и корреляции континентальных отложений. Листовая флора и спорово-пыльцевой анализ дают возможность выделять стратиграфические подразделения вплоть до подъярусов и зон. Кроме того, водоросли и высшие растения, обитающие в водной среде, позволяют установить температуру бассейна (холодноводные, тепловодные), его глубину и соленость. Ископаемые наземные растения помогают реконструировать климатические пояса, проводить палеофлористическое районирование суши и т.д.
Породообразующая роль растений значительна, а для образования торфа, горючих сланцев и углей — исключительна. В процессе углеобразования участвовали высшие растения, обитавшие преимущественно во влажных тропиках и создававшие заросли типа мангровых. Наиболее интенсивное углеобразование происходило в карбоне-перми и юре, в гораздо меньшей степени — в девоне и палеогене. В настоящее время продолжается массовое накопление органического вещества в манграх и болотах различных широт (торфяники). Образование углей в карбоне происходило за счет плауновидных, хвощевидных и папоротниковидных; в юре — папоротниковидных и пинофитовых; в палеогене — папоротниковидных, пинофитовых и цветковых; в настоящее время — цветковых и листостебельных мхов (сфагнум). В создании биогенных карбонатных пород (особенно известняков) принимают участие различные водоросли: золотистые, красные, зеленые, харовые. Диатомовые водоросли образуют кремневые породы (диатомиты, трепела, опоки). Горючие сланцы ордовика — кукерситы — состоят из массовых скоплений органических оболочек рода Gloeocapsomorpha, видимо, принадлежавших зеленым водорослям. Горючие сланцы в различные период могли образовываться за счет скопления другой органики, как, например, доманиковые сланцы позднего девона.