

Отдел харовые водоросли. Divisio charophyta
(греч. chara — дикая капуста, полевой тмин; phyton — растение)
Харовые водоросли — многоклеточные организмы, близкие к. зеленым водорослям, с которыми их иногда объединяют. Прямостоящие слоевища харовых имеют признаки сходства с высши- ми растениями. Так, членистые слоевища, мутовчатые ответвления («листья») напоминают собой хвощи (рис. 35). Размножение вегетативное или половое. Харовые живут в пресных водоемах либо опресненных лагунах, нередко образуя обширные заросли. Известно 6 родов и около 300 видов. Размеры от 20-30 см до 1-2 м.
Харовые появились в позднем силуре и существуют поныне. В ископаемом состоянии нередко сохраняются вместилища яйцеклеток — оогонии. Массовые скопления обызвествленных оого- ний создают породы, называемые «харовый туф», или «хароцит».
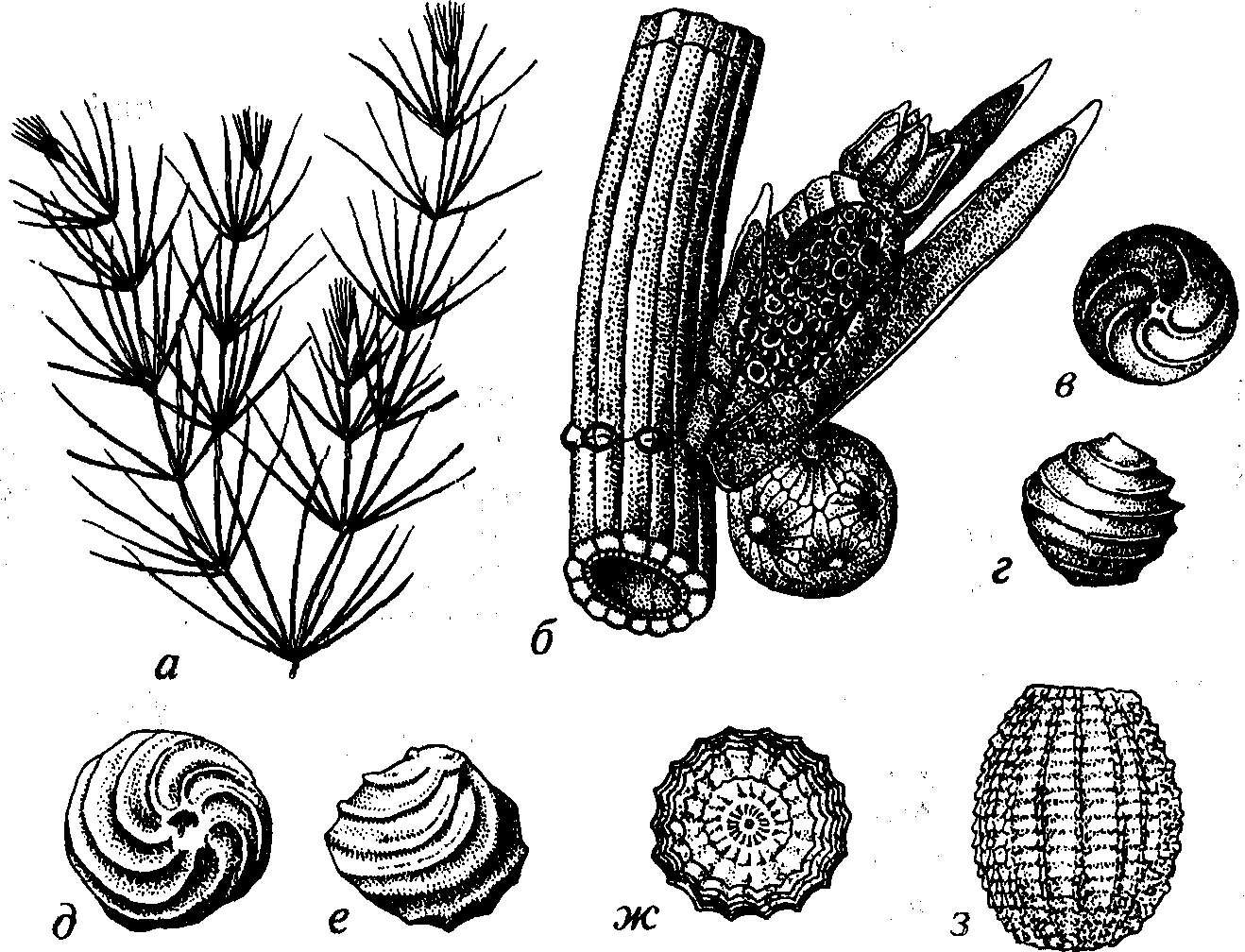
Рис. 35. Отдел Charophyta (S2-Q) а-г — Chara (P2-Q): a — слоевище, б — часть слоевища, вверху оогоний и вниз;; антеридий, в, г — оогонии; д, е — Trochiliscus (D), оогонии; ж, з — Sycidium, оогонии (O2-С)
ПОДЦАРСТВО ВЫСШИЕ РАСТЕНИЯ.
SUBREGNUM TELOMOPHYTA
(греч., telos — конец)
Подцарство высших растений отличается от полцарства низших растений следующими особенностями: 1. Тело расчленено на корень, стебель, листья и органы размножения. 2. Специализация клеток приводит к образованию различных специфических тканей, осуществляющих проводящую, защитную, механическую и другие функции. 3. Среда обитания наземная, хотя имеются некоторые вторично-водные формы. 4. Закономерное чередова- ние полового (гаметофит) и бесполого (спорофит) поколений. При этом в жизненном цикле всех высших растений (кроме мохообразных) резко преобладает длительность существования спорофита над гаметофитом, особенно у древесных форм. Основным звеном в эволюции высших растений явились преобразование и усложнение органов размножения с сокращением и последующей редукцией гаметофита как самостоятельного растения.
Выход растений на сушу и освоение наземной среды сопровождался усложнением общего плана строения и совершенствованием проводящей системы. Наиболее примитивные высшие растения имели стебель и просто устроенные органы размножения, на следующем этапе развития появились листья и корни.
Строение и функции основных частей растения в общем плане достаточно известны. Стебель — осевая часть растения — выполняет две основные функции: опорную и проводящую, а также несет побеги, листья, органы размножения. С помощью листьев происходят фотосинтез, газообмен и испарение воды. Корень является подземной частью растения, закрепляет его на субстрате, снабжает водой и минеральными солями.
Ткани растений можно подразделить на 5 различных типов: покровные, механические, проводящие, основные и образовательные; последний тип иногда рассматривается как разновидность основной ткани. Первые три типа тканей четко обособлены, а четвертый тип — основная ткань — заполняет все оставшееся пространство между ними.
Покровная ткань (эпидермис, эпидерма, кожица) защищает растение от механических повреждений, а также от излишнего испарения и перегревания. Клетки эпидермиса снаружи обволакивает слой воскоподобного вещества — кутина, а образуемая им пленка называется кутикулой. Газообмен и испарение влаги осуществляются через устьица — отверстия в эпидермисе, которые с двух сторон ограничены двумя клетками дуговидной формы (см. рис. 62).
Механические ткани состоят из вытянутых, нередко волокнистых клеток, которые обеспечивают сопротивление механическим воздействиям (например, изгибание ствола дерева) и способ- ствуют прочности растения в целом. Механические ткани растений используются для изготовления хлопчатобумажных нитей и тканей, веревок и канатов.
Проводящие ткани обеспечивают встречную транспортировку снизу вверх от корней к листьям и сверху вниз от листьев к корням. Группировка клеток, выполняющая функцию перемещения вверх воды с минеральными солями, получила название ксилема (греч. xilon — древесина, срубленное дерево). Она сложена удлиненными клетками — трахеидами — со скошенными концами. Длина трахеид колеблется от 1 до 5 мм. Трахеиды имеют разнообразную скульптуру: кольчатую, спиральную, лестничную, сетчатую. Скульптура возникает за счет чередования утолщенных и неутолщенных участков стенки трахеид. Лишь неутолщенные участки стенки могут пропускать воду.
У цветковых растений трахеиды заменяются сосудами, возникшими в результате срастания нескольких клеток и исчезновения перегородок между ними. Сосуды в десятки и сотни раз длиннее трахеид. Трахеиды и сосуды теряют цитоплазму и поэтому утрачивают способность к делению.
Движение органических продуктов фотосинтеза от листьев к корням осуществляется у высших растений с помощью удлиненных ситовидных трубок со скошенными концами. Ситовидные трубки образуют флоэму.
Проводящие ткани (ксилема и флоэма) соседствуют друг с другом или на границе между ними располагается камбий. В сочетании с механической тканью они формируют проводящие пучки, которые по аналогии с животными можно рассматривать как «артерии и вены» растений, соединяющие все органы живого организма. Проводящие пучки осевой части стебля и корня, ограниченные первичной корой (эпидерма), образуют стелу (греч. stele — столб).
Различные комбинации расположения ксилемы и флоэмы в проводящих пучках позволяют выделить несколько различных типов стел (рис. 36).
У древних высших растений в осевой части стебля находится цилиндр ксилемы, ее окружает кольцо флоэмы, при этом образуется единый центральный проводящий пучок — протостела (греч. protos — первый). Протостелический тип стелы характерен для риниофитов, некоторых папоротников и для корней всех высших растений.
От протостелы возникли другие типы стел и в первую очередь актиностела и сифоностела (эктофлойная и амфифлойная), Актиностела (греч. aktis, aktinos — луч + стела) представляет со- бой разновидность протостелы, характеризующуюся звездчатой формой в поперечном сечении. Такой тип стелы известен у некоторых плауновидных.
Оба типа сифоностелы в отличие от протостелы имеют в осевой части стебля сердцевину, заполненную основной тканью с рыхлым расположением клеток. Основная ткань сердцевины из-за рыхлой структуры может подвергаться быстрому разрушению, и иногда центральная часть стебля становится полой (например, некоторые злаки). Эктофлойная сифоностела (греч. ektos — вне, снаружи + флоэма) имеет в центре сердцевину, окруженную кольцом ксилемы, за которым следует кольцо флоэмы. Флоэма по отношению к ксилеме располагается снаружи. Амфифлойная сифоностела (греч. amphi — двойной с обеих сторон) характеризуется тем, что кольцо ксилемы окружено двумя кольцами флоэмы:наружным и внутренним. Так устроена стела лепидодендроновых и некоторых папоротников.
Распадение сифоностелы как единого осевого цилиндра привело к образованию еще нескольких типов стел. Основные из них: диктиостела, эвстела, артростела, атактостела. При распадении амфифлойной сифоностелы на отдельные пучки (меристелы) формируется диктиостела, причем флоэма в меристелах образует замкнутое кольцо вокруг ксилемы. В результате формируется закрытый проводящий пучок, лишенный возможности роста в тол- щину. Диктиостела встречается преимущественно у папоротников.
Аналогичный процесс распадения эктофлойной сифоностелы на отдельные пучки, разделенные радиальными лучами основной ткани, привел к возникновению эвстелы (греч. ей — хорошо, настоящий + стела). Однако в отличие от диктиостелы проводящие пучки эвстелы являются открытыми, между ксилемой и флоэмой имеется камбий, и поэтому возможен вторичный рост в толщину. Эвстела встречается у некоторых папоротниковидных, у большинства голосеменных и у двудольных покрытосеменных.
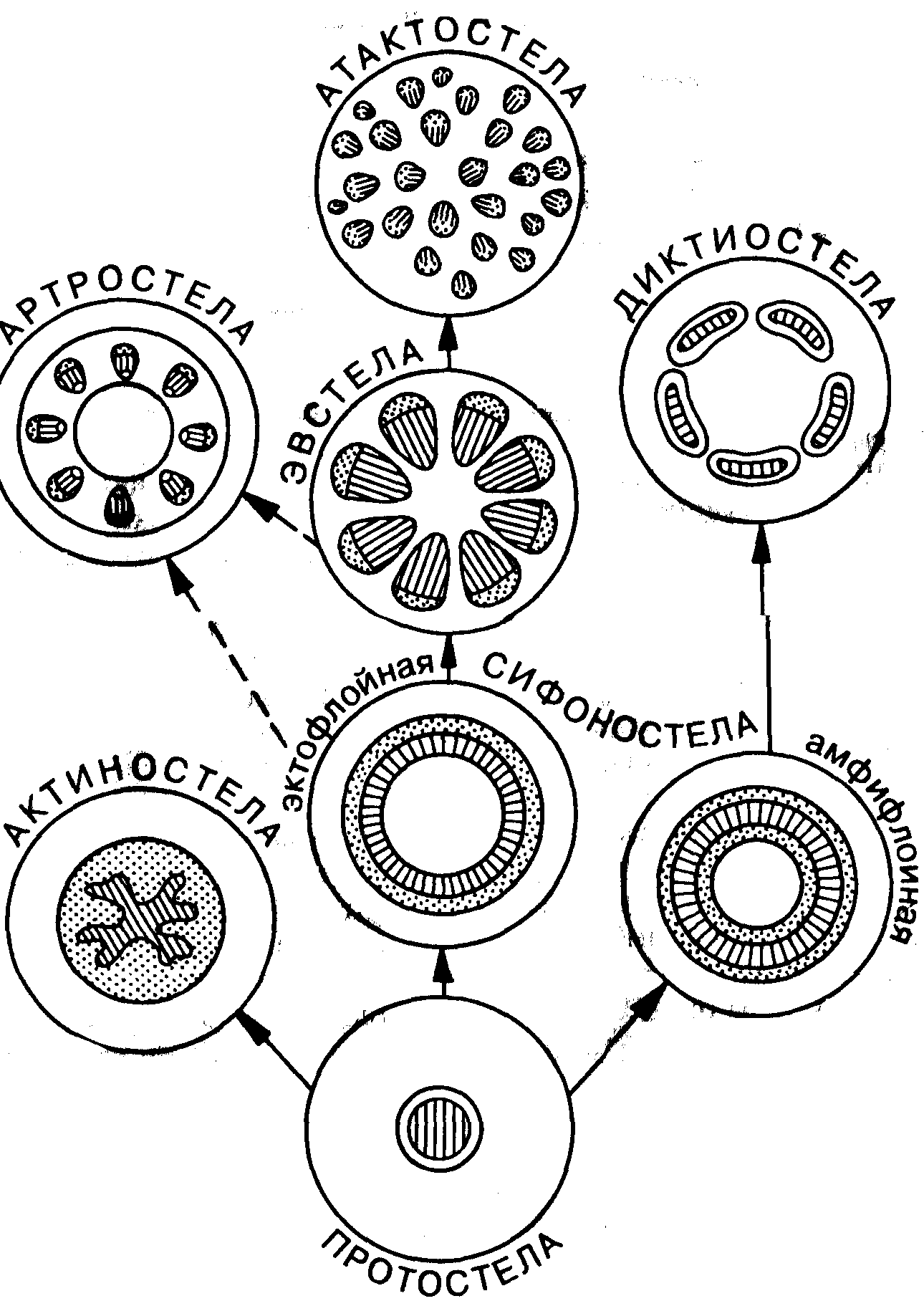
Рис. 36. Основные типы.стел высших растений (Тахтаджии, 1954)
От эктофлойной сифоностелы, а возможно, от эвстелы возникла характерная для хвощей артростела (греч. arthron — сустав + стела), представляющая собой изолированные проводящие пучки. Вокруг пучков артростелы располагается внутренняя кора.
Последний тип стелы — атактостела (греч. ataktos — беспорядочный + стела) отличается от эвстелы и артростелы рассеянны расположением изолированных проводящих пучков закрытого типа. Атактостела характерна для однодольных покрытосеменных растений. Так как многие однодольные имеют полую сердцевину, то у них проводящие пучки располагаются только по периферии.
Основная ткань заполняет пространство между названными видами тканей. Она состоит из клеток, имеющих по всем направлениям равные размеры и способных изменять свои функции. Главное свойство основной ткани — способность синтезировать и запасать органические вещества.
Образовательная ткань — меристема (греч. meristos — делимый) состоит из клеток, способных делиться и давать другие типы тканей. Увеличение стебля в высоту и удлинение корня происходят за счет верхушечных конусов нарастания (верхушечные меристемы). Это первичный рост. Утолщение стебля и «вторичный» рост являются функцией камбия. Последний представляет собой один ряд клеток образовательной ткани (цилиндрическая меристема), который располагается в проводящем пучке на границе ксилемы и флоэмы.
Для высших растений характерны листья (рис. 37). В широком понимании листьями называют плоские боковые выросты, идущие от побега. Они имеют различное происхождение и анатомическое строение. Выделяют три группы листьев: филлоидные (греч. phyllon — лист, пластинка), настоящие и филлидные. У риниофитов шиловидные выросты эпидермиса стебля не имеют внутри проводящих пучков; их называют эмергенцами (лат. emergens — выбивающийся). Листовидные выросты мхов назвали филлидными листьями, или филлидами. У настоящих листьев проводящие пучки отходят от осей
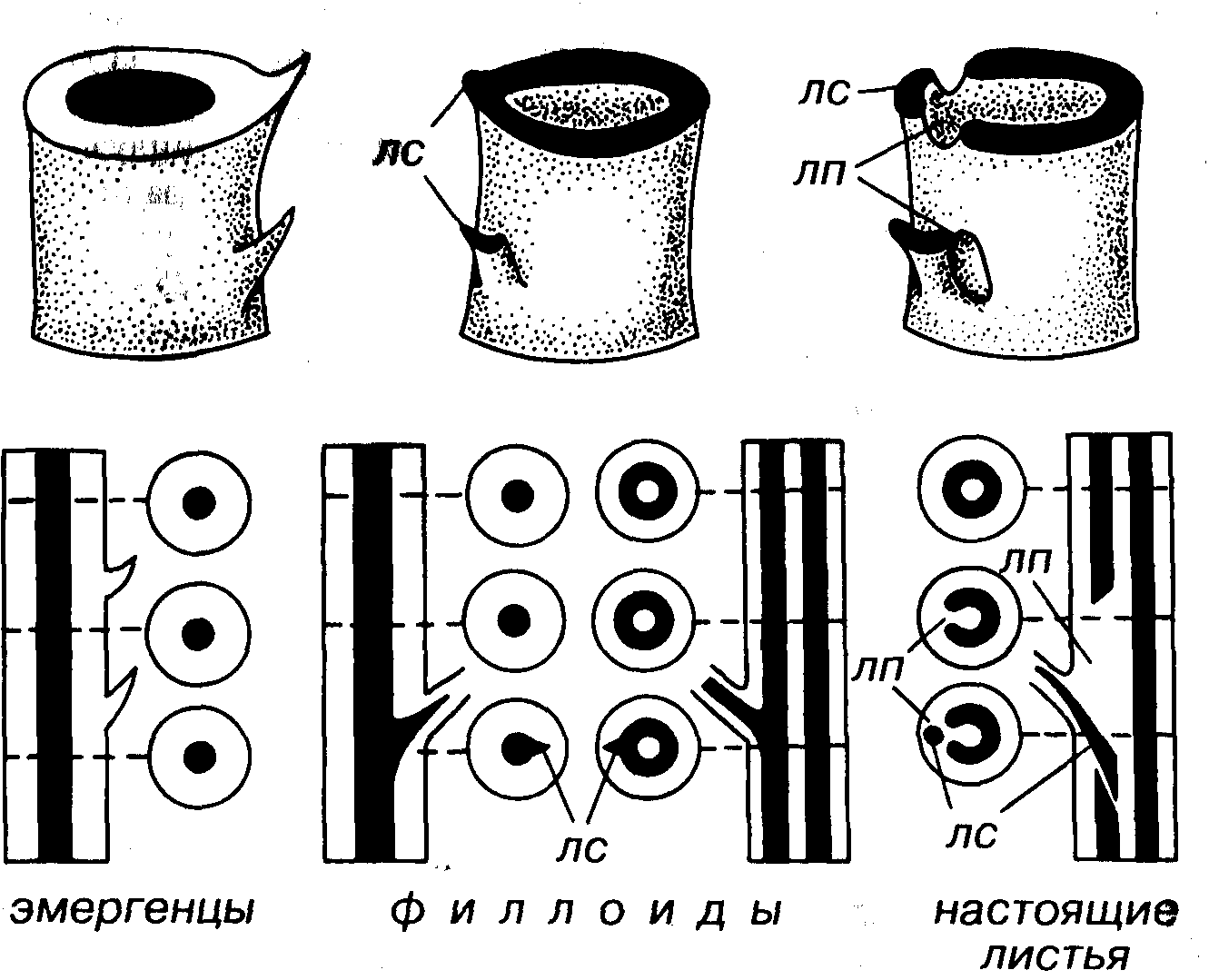
Рис. 37. Схема строения листьев и листоподобных образований Верхний ряд — объемные реконструкции, нижний ряд — продольные и попереч- ные сечения, лп — листовой прорыв, лс — листовой след, проводящая система
зачернена
стелы, прорывая ее в виде щелевидно-оконтуренного пространства (листовой прорыв), и проходят в лист, оставляя листовой след. Филлоидные листья, или филлоиды, являются выростами только поверхностных тканей. В такие листья заходит проводящий пучок, имеется листовой след, но отсутствует листовой прорыв. Филлоиды характерны для плауновидных, а настоящие листья — для хвощевидных, папоротниковидных, голосеменных и покрытосеменных, возможно, и для некоторых плауновидных.
Как уже говорилось, особенностью высших растений является закономерное чередование полового (гаметофит) и бесполого - (спорофит) поколений при явном преобладании одного из них. Гаметофит обладает одинарным (гаплоидным), а спорофит — двойным (диплоидным) набором хромосом.
Рассмотрим жизненный цикл папоротника (рис. 38). Хорошо знакомое растение представляет собой бесполое поколение — спорофит. На нижней стороне крупных листьев папоротника нередко можно наблюдать группы спорангиев (органы бесполого размножения), в которых в результате редукционного деления созревают споры с гаплоидным набором хромосом. Спорангии иногда группируются в сорусы или срастаются вместе, давая синангии. Спо- рангии могут быть защищены особым образованием — индузием. Споры в большинстве случаев покрыты плотной защитной оболочкой, которая нередко несет разнообразную скульптуру. Форма спор различна, их размеры изменяются от 3-5 до 25-50 мкм. Споры высших растений в отличие от таковых низших не имеют жгутиков; при созревании, прежде чем попасть в почву, они могут быть перенесены ветром или водой иногда на значительные расстояния. Высшие растения могут продуцировать однотипные споры (равноспоровые) или разнотипные (разноспоровые: микро- и мегаспоры).
Каждая спора представляет собой специализированную клетку, участвующую в процессе полового размножения. Гаплоидные споры дают начало новому — половому — поколению (гаметофиту), производящему половые продукты. Гаметофит обычного папоротника представляет собой маленькую пластинку сердцевидной формы (заросток) размером от долей миллиметра до 3 см. На нижней стороне заростка формируются археогонии — женские половые органы, в которых развиваются яйцеклетки, и антеридии — мужские половые органы, в которых созревают сперматозоиды. Этот заросток является обоеполым, но у других растений встречаются и разнополые заростки. Оплодотворенная яйцеклетка преобразуется в диплоидную зиготу (греч. zygote — соединенная в пару), давая начало будущему диплоидному спорофиту. При этом происходило усложнение процесса полового размножения. Микроспоры постепенно преобразовались в пыльцу, а мегаспорангии с мегаспорами — в семязачатки (= семяпочки). После оплодотворения семяпочки пыльцой возникает семя.
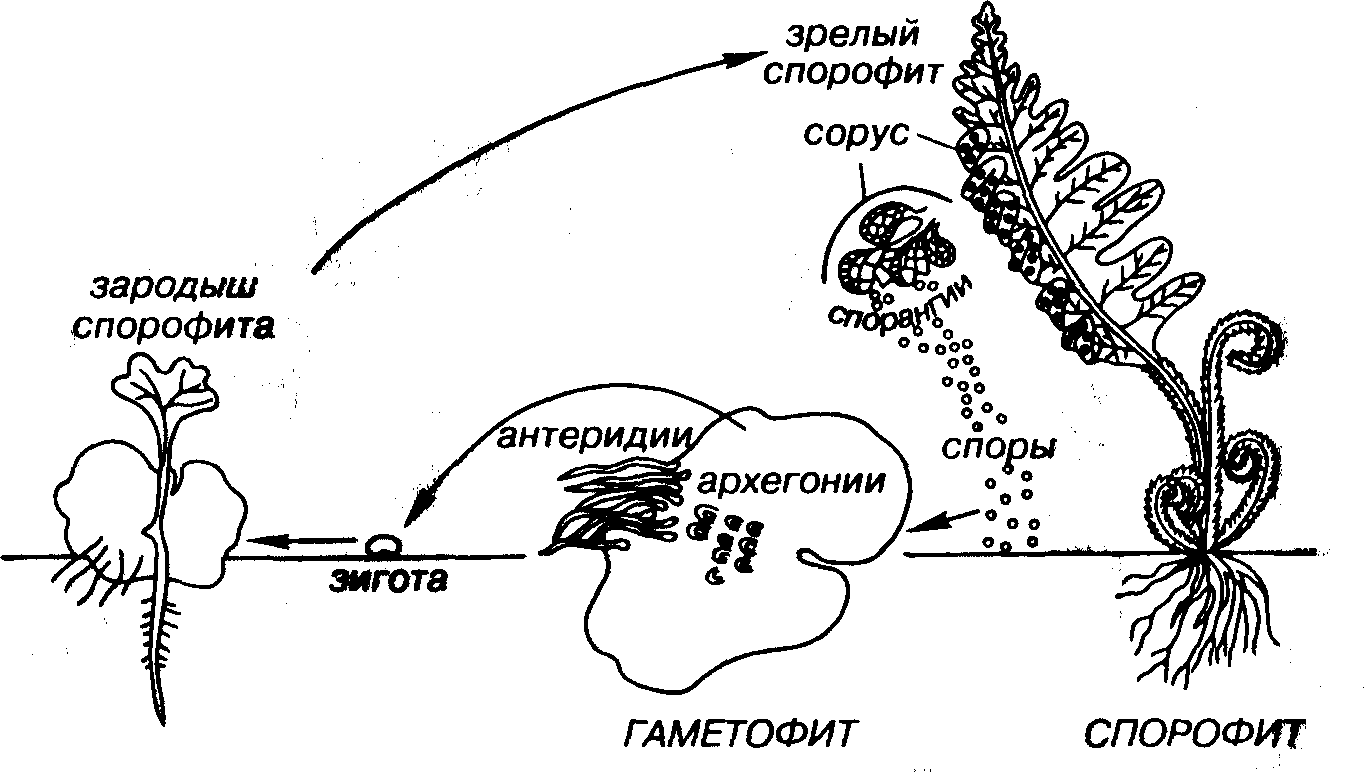
Рис. 38. Чередование поколений у современного папоротника
В соответствии со способом размножения подцарство высших растений разделено на два надотдела: Sporophyta (споровые) и Spermatophyta (семенные).