

естественно
.PDF221
вождался сменой выраженной левосторонней латерализации амплитуд бета-2-, бета-1-, альфа- и тета-ритмов правосторонней
иувеличением левосторонней латерализации амплитуды дель- та-ритма. Полученные результаты в определенной степени согласуются с представлениями о возможности смены доминантности полушарий в разные фазы сна и об относительном повышении активности правого полушария во время быстрого
илевого – во время медленного сна.
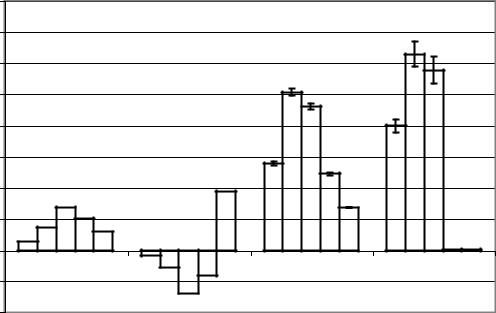
Переход от десинхронизации к синхронизации при отведении от препарата ИПМ сопровождался увеличением левосторонней латерализации амплитуд бета-2-, бета-1- и альфаритмов и уменьшением левосторонней латерализации амплитуд тета- и дельта-ритмов (рис. 50).
С учетом описанных выше данных можно высказать предположение, что структуры ствола (в частности, РФ) и мозжечка наряду с собственно корковыми механизмами в значительной степени обеспечивают управление ФМПА в условиях интактности мозга.
Следует подчеркнуть, что показатели ФПМА в условиях ИПМ не только были положительными, т.е. проявлялась от четливая латерализация электрической активности в левое полушарие, но и при этом их модули в два–пять раз превышали аналогичные величины, характерные для ИМ. Указанная особенность, возможно, связана с тем обстоятельством, что связи полушарий со срединными образованиями мозга разного уровня организованы различным образом. Кора левого полушария имеет более развитые связи со стволовыми структурами, тогда как правого – с диэнцефальными (Брагина Н. Н., Доброхотова Т. А., 1988).
Полученные результаты также могут свидетельствовать о том, что влияния из ствола и мозжечка в некоторой степени нивелируют собственную ФМПА полушарий; такое заключение согласуется с представлениями о генерализованном преобладании ингибиторных процессов в левом полушарии, а активационных – в правом.
222
%
40
35
30
25
20
15
10
5
0
-5
-10
int_desin |
int_sin |
si_des |
si_sin |
|
группы опытов - ритмы ЭКоГ |
|
|

 B2A
B2A  B1A
B1A 
 AA
AA 
 TA
TA 
 DA
DA
Рис. 50. Диаграммы коэффициентов функциональной межполушарной асимметрии амплитуд ритмов 2, 1, и ЭКоГ, отводимой от интактного мозга и препарата изолированного мозга, int_desin – интактный десинхронизация, int_sin – интактный синхронизация, si_desin – препарат изолированного переднего мозга десинхронизация, si_sin - препарат изолированного переднего мозга синхронизация.
3.7.5.Промежуточный мозг
Кпромежуточному мозгу относятся:
1)зрительный мозг (thalamencephalon);
2)подбугровая (субталамическая) часть, гипоталамус (hypothalamus).
В зрительном мозге различают:
а) зрительный бугор (thalamus opticus),
б) надбугровую область (epithalamus),
в) забугровую область (metathalamus). 3.7.5.1. Таламус
223
Зрительный бугор имеет яйцевидную форму. Задний полюс зрительного бугра утолщен и слегка вытянут, его называют подушкой (pulvinar thalamus). В зрительных буграх находятся многочисленные, свыше 120 ядерных образований, в которые поступает афферентация из различных отделов головного мозга (коры головного мозга, мозжечка, центров экстрапирамидной системы, миндалевидного комплекса).
Зрительный бугор - это коллектор всех видов чувствительности. Уолкер, один из специалистов по функциональной организации зрительного бугра, писал: "Таламус является посредником, в котором сходятся все раздражения от внешнего мира и, видоизменяясь здесь, направляются к подкорковым и корковым центрам таким образом, чтобы организм смог адекватно приспособиться к постоянно меняющейся среде. Таламус, как видно, таит в себе тайну того, что происходит в коре головного мозга".
Зрительный бугор является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом (подтверждением тому служит наличие в таламусе около 120 разнофункциональных ядер). Ядра образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на 3 группы: передняя проецирует аксоны своих нейронов в поясную извилину коры большого мозга; медиальная — в лобную долю коры; латеральная — в теменную, височную, затылочную доли коры. По проекциям определяется и функция ядер. Такое деление не абсолютно, так как одна часть волокон от ядер таламуса идет в строго ограниченные корковые образования, другая — в разные области коры большого мозга.
1. Предложено несколько классификаций ядер зрительных бугров, в основу которых положены различные принципы. Уолкер (1938) разделил ядра таламуса на шесть групп.
В 1927 году Хасслер на основании характера связей ядер зрительных бугров с корой головного мозга выделил две системы ядер:
1. Паллиоталамические ядра. К ним относятся ядра, имеющие двусторонние связи с корой головного мозга (передняя
224
ядерная группа, медиальная ядерная группа, латеральная ядерная область, медиальные коленчатыетела, латеральные коленчатые тела, подушка).
2.Трункоталамические ядра. Эти ядра не имеют прямых связей с корой головного мозга (центральное, или перивентрикулярное, серое вещество таламуса, интраламинарная группа и ретикулярная зона).
В 1976 году Адрианов на основании эволюционных и физиологических признаков выделил три группы ядер:
1.Диффузная таламическая система (неспецифические ядра). Данная группа ядер представляет филогенетически древнюю часть таламуса и состоит из интраламинарной группы ядер средней линии. Диффузная таламическая система характеризуется следующими признаками:
а) получает афференты из филогенетически древней экстралемнисковой системы, спинного мозга, бульбарных отделов ретикулярной формации;
б) не имеет прямого выхода к коре головного мозга. Выход в кору мозга осуществляется через ретикулярное ядро таламуса.
2.Группа специфических (релейных) ядер. Данная группа подразделяется на сенсорные и несенсорные релейные ядра.
Релейные ядра:
а) получают афференты от лемнисковых систем (спинальная, тригеминальная, слуховая, зрительная), от структур мозжечка, гипоталамуса, полосатого тела;
б) имеют прямой выход в кору головного мозга; в) характеризуются топографической и функциональной
разграниченностью проекций к определенным областям коры головного мозга;
г) имеют соматотопическую организацию.
3.Ассоциативные ядра таламуса. Данная группа ядер характеризуется следующими признаками:
а) не имеет прямого афферентного входа из периферии; б) афференты этих ядер берут начало в других ядрах тала-
муса;
в) имеется развитая система связей данной группы ядер с ассоциативными полями коры головного мозга;
225
г) данная группа ядер осуществляет наиболее сложные интегративные процессы по сравнению с другими ядрами таламуса.
Как отмечено выше, таламус является коллектором всех видов чувствительности. Это объясняется тем, что в таламус входит 12 различных афферентных систем:
а) зрительная афферентная система; б) слуховая афферентная система; в) вкусовой тракт; г) спинно-таламический тракт; д) спинно-кортикальный тракт; е) спинно-цервико-таламический тракт; ж) дорсальный спинно-церебеллярный тракт; з) вестибулоталамический тракт; и) церебелло-таламический тракт; к) пучок лентикулярной петли; л) внутренний лапидарный тракт; м) наружный палидарный тракт.
Латеральное коленчатое тело имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
В медиальное коленчатое тело (МТК) поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферетные пути от медиальных коленчатых тел идут в височную зону коры большого мозга, достигая там первичной слуховой области коры. МКТ имеет четкую тонотопичность. Следовательно, уже на уровне таламуса обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Многочисленность афферентных входов зрительных бугров приводит к тому, что любой афферентный импульс, прежде чем поступит в кору головного мозга, непременно достигнет зрительных бугров. При этом следует подчеркнуть, что зрительные бугры не являются только проводниками афферентации в кору мозга, в зрительных буграх совершаются сложные интегративные процессы. Только после соответствующей трансформации на уровне зрительных бугров афферентация поступает в кору головного мозга.
Основной эфферентный выход из таламуса представлен сис-
226
темой волокон, которая проводит афферентацию из зрительных бугров в кору мозга.
В основе интегративной деятельности таламуса лежит многоуровневый принцип организации интегративных процессов.
Первый уровень интеграции - это взаимодействие процес-
сов на синаптическом уровне.
Второй уровень интеграции - это интрануклеарный уро-
вень, выражающийся в объединении значительной группы нейронов таламического ядра с помощью внутриядерных тормозных интернейронов. Тормозный интернейрон устанавливает связи со многими релейными нейронами. Возбуждение такого интернейрона вызывает торможение значительной группы релейных нейронов, что приводит к синхронизации их деятельности. Такая деятельность обеспечивает контроль входа в таламические ядра и контроль выхода афферентации из таламуса (интраталамический уровень). Интрануклеарные и интраталамические интегративные процессы имеют большое значение в осуществлении специфических функций определенных ядер таламуса.
Третий уровень интеграции - это таламокортикальный уровень интеграции, являющийся наиболее сложным. Этот уровень интеграции контролирует сложнейшие формы моторного, эмоционального поведения; формирует болевые ощущения; осуществляет регуляцию многообразных вегетативных функций (дыхания, пищеварения, деятельности сердца, тонуса сосудов и др.). За счет связей нейронов таламуса с различными участками коры головного мозга и связей коры мозга с нейронами таламуса формируется сложнейшая система таламокортикальных взаимоотношений, в которой зрительный бугор выступает как ближайший помощник коры головного мозга в регуляции моторных и соматических функций, а также сложных форм поведения.
Сложное строение таламуса, наличие в нем взаимосвязанных специфических, неспецифических и ассоциативных ядер, позволяет ему организовывать такие двигательные реакции, как сосание, жевание, глотание, смех. Двигательные реакции интегрируются в таламусе с вегетативными процессами, обеспечивающими эти движения.
227
Конвергенцию сенсорных стимулов в таламус обусловливает возникновение так называемых таламических и болей, которые возникают при патологических процессах в самом таламусе.
3.7.5.2. Гипоталамус
Гипоталамус (hypothalamus, подбугорье) — структура промежуточного мозга, входящая в лимбическую систему, организующая эмоциональные, поведенческие, гомеостатические реакции организма.
Морфофункциональная организация. Гипоталамус имеет большое число нервных связей с корой большого мозга, подкорковыми узлами, зрительным бугром, средним мозгом, мостом, продолговатым и спинным мозгом.
В состав гипоталамуса входят серый бугор (tuber cinereum), воронка (infundibulum) с нейрогипофизом и сосцевидные тела (corpora mamillaria). Морфологически в нейронных структурах гипоталамуса можно выделить около 50 пар ядер, имеющих свою специфическую функцию. Топографически эти ядра можно объединить в 5 групп: 1) преоптическая группа имеет выраженные связи с конечным мозгом и делится на медиальное и латеральное предоптические ядра; 2) передняя группа, в состав которой входят супраоптическое, паравентрикулярные ядра; 3) средняя группа состоит из нижнемедиального и верхнемедиального ядер; 4) наружная группа включает в себя латеральное гипоталамическое поле и серобугорные ядра; 5) задняя группа сформирована из медиальных и латеральных ядер сосцевидных тел и заднего гипоталамического ядра.
Ядра гипоталамуса имеют мощное кровоснабжение, подтверждением чему служит тот факт, что ряд ядер гипоталамуса обладает изолированным дублирующим кровоснабжением из сосудов артериального круга большого мозга (виллизиев круг). На 1 мм2 площади гипоталамуса приходится до 2600 капилляров, в то время как на той же площади V слоя предцентральной извилины (моторной коры) их 440, в гиппокампе — 350, в бледном шаре — 550, в затылочной доле коры большого мозга (зрительной коре) — 900. Капилляры гипоталамуса высоко-
228
проницаемы для крупномолекулярных белковых соединений, к которым относятся нуклеопротеиды, что объясняет высокую чувствительность гипоталамуса к нейровирусным инфекциям, интоксикациям, гуморальным сдвигам.
У человека гипоталамус окончательно созревает к возрасту 13— 14 лет, когда заканчивается формирование гипота- ламо-гипофизарных нейросекреторных связей. За счет мощных афферентных связей с обонятельным мозгом, базальными ганглиями, таламусом, гиппокампом, корой большого мозга гипоталамус получает информацию о состоянии практически всех структур мозга. В то же время гипоталамус посылает информацию к таламусу, ретикулярной формации, вегетативным центрам ствола мозга и спинного мозга.
Нейроны гипоталамуса имеют особенности, которые и определяют специфику функций самого гипоталамуса. К этим особенностям относятся чувствительность нейронов к составу омывающей их крови, отсутствие гематоэнцефалического барьера между нейронами и кровью, способность нейронов к нейросекреции пептидов, нейромедиаторов и др.
Роль гипоталамуса в регуляции вегетативных функций. Влияние на симпатическую и парасимпатическую регуляцию позволяет гипоталамусу воздействовать на вегетативные функции организма гуморальным и нервным путями.
Раздражение ядер передней группы сопровождается парасимпатическими эффектами. Раздражение ядер задней группы вызывает симпатические эффекты в работе органов. Стимуляция ядер средней группы приводит к снижению влияний симпатического отдела автономной нервной системы. Указанное распределение функций гипоталамуса не абсолютно. Все структуры гипоталамуса способны в разной степени вызывать симпатические и парасимпатические эффекты. Следовательно, между структурами гипоталамуса существуют функциональные взаимодополняющие, взаимокомпенсирующие отношения.
В целом за счет большого количества связей, полифункциональности структур гипоталамус выполняет интегрирующую функцию вегетативной, соматической и эндокринной регуляции, что проявляется и в организации его ядрами ряда конкретных функций. Так, в гипоталамусе располагаются центры
229
гомеостаза, теплорегуляции, голода и насыщения, жажды и ее удовлетворения, полового поведения, страха, ярости, регуляции цикла бодрствование—сон. Все эти центры реализуют свои функции путем активации или торможения автономного (вегетативного) отдела нервной системы, эндокринной системы, структур ствола и переднего мозга.
Впервые в 1928 году Шаррер открыл способность нейронов супраоптического и паравентрикулярного ядер переднего отдела гипоталамуса к "секреции" веществ нейрогормональной природы: антидиуретического гормона (АДГ, вазопрессин) и окситоцина. Это открытие привело к установлению эндокринных свойств гипоталамуса. Формируясь в нейронах этих ядер, гормоны в виде гранул транспортируются по их аксонам в заднюю долю гипофиза (нейрогипофиз). Антидиуретический гормон регулирует обратное всасывание воды в почечных канальцах и воздействует на гладкую мускулатуру артериол, что приводит к повышению артериального давления. Окситоцин специфически действует на гладкую мускулатуру матки и молочных желез, стимулируя соответственно сокращение матки и секрецию молока молочными железами.
Супрахиазматическое (супрахиазменное) ядро расположено над зрительным перекрестом в основании гипоталамуса (рис. 51). Оно получает вход от зрительной системы и отвечает за восприятие светлого времени суток как времени бодрствования, а также поддерживает суточный ритм.
«Биологические часы» человека расположены на уровне супрахиазмальных ядер гипоталамуса. Они имеют периодичность хода, определяемую скоростью определенных биохимических реакций в клетке, составляющую около 25 часов (у большинства людей), и поэтому нуждаются в ежедневной "подстройке".
Настройка «биологических часов» осуществляется с помощью солнечного света: свет возбуждает особую группу светочувствительных клеток сетчатки, содержащих специальный фотопигмент меланопсин.
Активация клеток супрахиазмальных ядер (СХЯ), содержащих два типа нейронов – выделяющих глутамат и выделяю-

230
щих нейропептид Y, так же как активация клеток латерального гипоталамуса (ЛГ), синтезирующих пептид гипокретин/орексин, и клеток медиальной префронтальной коры (ПК),
Рис. 51. Медиальная поверхность полушария.
выделяющих глутамат, может также непосредственно активировать обширные области коры мозга. Активация этих областей мозга вызывает прямо или косвенно (через таламокортикальные, гипоталамо-кортикальные и/или базалокортикальные восходящие пути) активацию коры, а также глобальную активацию всего мозга, необходимую для запуска и поддержания состояния поведенческого бодрствования (Datta S., MacLean R.R., 2007;Ковальзон В.М., 2011).
Тесное функционирование гипофиза с гипоталамусом позволяет рассматривать данные образования как единый структурно-функциональный комплекс, который называют "эндокринным мозгом".
Нейроны ядер срединной группы гипоталамуса продуцируют так называемые рилизинг-факторы (либерины) и ингибирующие факторы (статины), которые регулируют активность передней доли гипофиза — аденогипофиз. В нем образуются такие вещества, как соматотропный, тиреотропный и другие гормоны. Наличие такого набора пептидов в структурах гипоталамуса свидетельствует о присущей им нейросекреторной функции.