

естественно
.PDF251
ГАМК-ергические) нейроны, активируют небольшое количество эффекторных нейронов, которые через возвратные коллатерали возбуждают тормозные интернейроны. Очевидно, что усиление дофаминергических влияний будет способствовать развитию тормозных эффектов, ослабление же или выключение этих влияний вызовет состояние гиперактивности стриариых нейронов. С этим заключением согласуются результаты специального анализа вопроса о тормозных и возбуждающих эффектах дофамина. Вместе с тем наличие двух видов дофаминовых рецепторов, активация которых дает тормозящий или возбуждающий эффект, свидетельствует о сложных функциональных отношениях, индуцируемых дофамином. Возможно, патогенетическая роль вовлечения разных дофаминовых рецепторов в разных патологических процессах различна.
Серотонинергичеекий аппарат. В хвостатых ядрах обнаружено высокое содержание соротонина, выявлены серотониновые терминали. Поскольку серотонинсодержащие нейроны в ядрах не обнаружены, указанные выше данные позволили сделать заключение о наличии серотонинергического входа в неостриатум. В опытах на крысах показано, что основной серотонинергический вход составляют волокна, идущие из дорсального ядра шва. Они восходят к неостриатуму в составе медиального пучка.
Как и дофаминовые, серотониновые волокна образуют сеть варикозных расширений, частично контактирующих на нейронах хвостатого ядра. Стимуляция ядер шва вызывает высвобождение серотонина, а коагуляция — снижение концентрации серотонина и ферментов, участвующих в его синтезе в хвостатых ядрах.
Активация серотонинового входа тормозит вызванный потенциал в неостриатуме при раздражении таламуса.
При электрофоретическом подведении серотонина у большинства нейронов возникает подавление спонтанной ритмики. ЭС дорсального ядра шва вызывает торможение подавляющего большинства нейронов. Часть нейронов (25— 33%) не дает реакции, небольшая часть реагирует возбуждением. Электрофоретическое подведение метсергида, блокато-
252
ра постсинаптического действия серотонина, устраняет тормозные эффекты раздражения ядра шва.
Таким образом, приведенные данные свидетельствуют о преимущественно тормозном действии серотонинергического входа на деятельность нейронов хвостатого ядра. Однако остается неясным функциональное значение этих тормозных эффектов, поскольку неясно, на каких нейронах они реализуются.
Гамкергический аппарат. Базальные ганглии и черная субстанция отличаются высоким содержанием ГАМК. ГАМК-ергические нейроны в ядре двух типов: мелкие вставочные нейроны с ветвящимися отростками внутри ядра и нейроны, аксоны которых формируют эфферентные пути. К ним относится стрионигральная ГАМК-ергическая система, обеспечивающая возвратную петлю с тормозным действием на дофаминергические нигростриарные нейроны. При удалении хвостатого ядра в черной субстанции повреждается до 50% ГАМК-ергических терминалей, снижается содержание ГАМК и ферментов его обмена. ЭС хвостатого ядра и электрофоретическое подведение ГАМК к дофаминергическим нейронам черной субстанции вызывает торможение большинства из этих нейронов. Фармакологическое нарушение дофаминовой передачи в хвостатом ядре приводит к ослаблению тормозного ГАМК-ергического действия на дофаминовые нейроны черной субстанции. Потенцирование выброса дофамина пресинаптическими терминалями или активация постсинаптических дофаминовых рецепторов в хвостатом ядре усиливает торможение дофаминергических нигростриарных нейронов, причем этот эффект может быть предотвращен повреждением стрионигрального пути. Имеются данные об участии ГАМК-ергических нейронов в формировании стриопаллидарных и паллидонигральных путей. Большинство эффекторных аксонов ГАМК-ергических нейронов, выходя из хвостатого ядра, дает возвратные коллатерали, регулирующие уровень активности нейронов ядра. Как уже указывалось, внутри ядра имеются ГАМКэргические интернейроны. Электрофоретическое подведение ГАМК к нейронам хвостатого ядра вызывает их торможе-
253
ние, которое может быть предотвращено антагонистами ГАМК. Высказано предположение, что возбуждающиеся при стимуляции черной субстанции нейроны неостриатума относятся к тормозным интернейронам, часть из них активируется опосредствованно через возвратные коллатерали эффекторных нейронов. Предполагается, что эти вставочные нейроны ГАМК-эргические.
Холинергический аппарат. В базальных ганглиях содержится значительное количество АХ. Выявлено несколько типов холинергических нейронов. Основную массу их представляют вставочные короткоаксонные нейроны типа II Гольджи, меньшая часть относится к эфферентным нейронам, формирующим проекционные системы (Hattori T. et al., 1976). Имеется также холинергический вход, заканчивающийся в головке хвостатого ядра и берущий начало в зоне инсерта и парафасцикулярных ядрах таламуса. Показано преимущественно возбуждающее действие АХ на нейроны ядра. По данным H. McLennan, D. H. York (1966), возбуж-
дающие эффекты под влиянием АХ возникают в нейронах, расположенных по наружному краю ядра, а тормозные — в нейронах центральной части. Представляет интерес своеобразие функциональных отношений между эффектами дофамина и АХ: нейроны, которые инги-бируются дофамином, возбуждаются АХ, и, наоборот, нейроны, которые возбуждаются дофамином, ингибируются АХ.
Норадреиергический аппарат. Содержание НА в хвостатых ядрах составляет 0,25—0,27 мг/г, что примерно в 25—30 раз меньше содержания дофамина в ядре. По содержанию НА хвостатые ядра уступают гипоталамусу и продолговатому мозгу, по в то же время уровень медиатора здесь выше, чем в коре больших полушарий и мозжечка. Содержание НА в хвостатых ядрах коррелирует с низкой активностью в этом ядре ключевого фермента — синтеза дофамин- |3-гидроксилазы. Неясен вопрос о происхождении стриарного НА. Указание на то, что дорсальный НА-пучок, который восходит от locus coeruleus и, проходя через неостриатум, возможно, образует сипаптические контакты с терминалями нигростриарных нейронов, не согласуется с ранее получен-
254
ными данными, согласно которым при повреждении locus coeruleus измененных НА-терминалей в хвостатом ядре обнаружено не было. Однако это не исключает наличия НАвхода в ядро из другого источника. Об этом свидетельствуют данные работы О. Lindvall, A. Bjorklund (1974), которые, используя высокочувствительный метод флюоресцентной микроскопии, обнаружили, что значительное количество НАаксонов, проходящих через хвостатое ядро, оканчиваются в нем.
Все изложенное подтверждает установившееся мнение, что хвостатые ядра играют весьма важную роль модулятора деятельности мозговых структур (Э. Б. Арушанян, В. А. Отеллин, 1976).
Базальные ганглии образуют многочисленные связи как между структурами, входящими в их состав, так и другими отделами мозга. Эти связи представлены в виде параллельных функциональных петель, связывающих кору больших полушарий (двигательную, соматосенсорную и лобную) с таламусом. Информация поступает из вышеперечисленных зон коры, проходит через базальные ядра (хвостатое ядро и скорлупу) и черное вещество в двигательные ядра таламуса, оттуда снова возвращается в эти же зоны коры — это скелетомоторная петля. Одна из таких петель управляет движениями лица и рта, контролирует такие параметры движения, как сила, амплитуда и направление.
Другая петля — глазодвигательная (окуломоторная) специализируется на регуляции движения глаз. Предполагается, что медиатором, возбуждающим кортикостриарные нейроны, является аминокислота — глутамат, а между базальными ганглиями и таламусом существуют в основном тормозные пути и их медиатором является ГАМК.
Многие авторы придерживаются мнения о том, что бледный шар находится под постоянным, сложившимся в эволюции тормозным контролем стриарного тела.
Если раздражать хвостатое ядро, то большая часть нейронов бледного шара тормозится, а меньшая возбуждается. В случае повреждения хвостатого ядра у животного появляется двигательная гиперактивность.
255
Взаимодействие черного вещества и хвостатого ядра основано на прямых и обратных связях между ними. Установлено, что стимуляция хвостатого ядра усиливает активность нейронов черного вещества. Стимуляция черного вещества приводит к увеличению, а разрушение — к уменьшению количества дофамина в хвостатом ядре. Установлено, что дофамин синтезируется в клетках черного вещества, а затем со скоростью 0,8 мм/ч транспортируется к синапсам нейронов хвостатого ядра. В хвостатом ядре в 1 г нервной ткани накапливается до 10 мкг дофамина, что в 6 раз больше, чем в других отделах переднего мозга, бледном шаре, в 19 раз больше, чем в мозжечке. Благодаря дофамину проявляется растормаживающий механизм взаимодействия хвостатого ядра и бледного шара.
При недостатке дофамина в хвостатом ядре (например, при дисфункции черного вещества) бледный шар растормаживается, активизирует спинно-стволовые системы, что приводит к двигательным нарушениям в виде ригидности мышц.
Хвостатое ядро и скорлупа связаны также со структурами, не входящими в эти петли: черной субстанцией, красным ядром, мозжечком, мотонейронами спинного мозга, Некоторые из этих структур, например черная субстанция, оказывают модулирующее влияние на функцию хвостатого ядра. В черной субстанции продуцируется дофамин, который транспортируется к нейронам хвостатого ядра и там накапливается. Высвобождаясь в хвостатом ядре, дофамин модулирует глютаматергическую кортикостриарную передачу информации, вызывая или ее облегчение, или торможение.
Полосатое тело (хвостатое ядро и скорлупа) принимают участие в организации и регуляции движений и обеспечении перехода одного вида движения в другое. Раздражение хвостатого ядра , с одной стороны, тормозит активность коры, подкорки, безусловные рефлексы (пищевой, оборонительный и др.) и выработку условных рефлексов. При поражении полосатого тела наблюдается ретроантероградная амнезия — выпадение памяти на события, предшествующие травме. Стимуляция хвостатого ядра тормозит восприятие зрительной, слуховой и других видов сенсорной информации. С другой стороны, хвостатое ядро оказывает возбуждающее действие. Так, при его поражении наблю-
256
дается ригидность мышц (повышение мышечного тонуса). Двустороннее повреждение полосатого тела побуждает к стремлению движения вперед, одностороннее — приводит к манежным движениям.
Скорлупа выполняет специфическую функцию: она отвечает за организацию пищевого поведения. При ее поражении наблюдаются трофические нарушения кожи, а ее раздражение вызывает слюноотделение и изменение дыхания.
Нами покзано противосудорожное действие хвостатого яд-
ра (рис. 57).
Функции бледного шара заключаются в провоцировании ориентировочной реакции, движения конечностей, пищевого поведения (жевание, глотание),
После разрушения бледного шара возникают гипомимия (маскообразное лицо), гиподинамия, эмоциональная тупость, тремор головы, конечностей при движении, монотонная речь.
При повреждениях бледного шара могут появиться подергивания отдельных мышц лица и туловища, нарушается синергизм движения конечностей при ходьбе.
Функции ограды мало изучены. Она имеет двусторонние связи с лобной, затылочной, височной корой, обонятельной луковицей, таламусом и другими базальными ядрами. Ограда оказывает облегчающее влияние на зрительные, слуховые и соматические раздражения. Атрофия ограды приводит к полной потере способности больного говорить, а ее раздражение вызывает моторные реакции со стороны пищеварительного тракта (жевание, глотание, рвотные движения), ориентировочную реакцию.
Таким образом, симптомы, связанные с нарушением двигательных функций при поражении базальных ганглиев, можно разделить на гипофункциональные, или недостаточность, и гиперфункциональные, или избыточность.
К первым относят акинезию (отсутствие движений), ко вторым — ригидность (повышение мышечного тонуса), баллизм (крупноразмашистый гиперкинез конечностей), атетоз («червеобразные»движения), хорею (быстрые подергивания), тремор (дрожание).
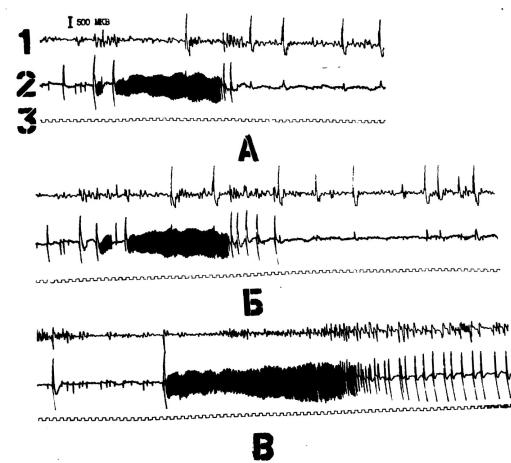
257
Рис. 57. Взаимоотношения между гиперсинхронной активностью в хвостатом ядре и судорожной активностью в круциатной извилине кошки.
Обозначения: А-после предварительных электростимуляций хвостатого ядра в круциатной извилине аппликацией 3,5% раствора пенициллина создан судорожный очаг. После введения в хвостатое ялро 900 ЕД пенициллина судорожная активность в круциатной извилине (зона 2) провоцировала возникновение гиперсинхронной активности в хвостатом ядре (зона 1), которая подавляла судорожную активность в зоне 2. Б-продолжение А, -после коагуляции зоны 1.
Обозначения: 1 – головка хвостатого ядра, 2 – круциатная извилина, 3 - отметка времени 1 с. Калибровка 500 мкв.
Поражение базальных ганглиев приводит к возникновению болезни Паркинсона, имеющей целый ряд симптомов, из которых главными являются ригидность, тремор и акинезия. Усилены тонические рефлексы растяжения, наблюдается восковая ригидность, сильное дрожание пальцев, губ и других частей тела.
258
Больному трудно начать и закончить движения, лицо его маскообразно, нарушена координация движений верхних и нижних конечностей во время ходьбы, он идет мелкими шажками, согнувшись вперед. При болезни Паркинсона нарушается планирование движений. Это заболевание связано с дегенерацией дофаминергических нейронов черного вещества, а результате в стриатуме резко падает содержание дофамина и происходит растормаживание холинергических нейронов, Поэтому лечение этого заболевания оказалось эффективным благодаря введению предшественника дофамина — L-дофа, так как сам дофамин не проходит через гематоэнцефалический барьер.
Хорея — наследственное дегенеративное заболевание базальных ганглиев, сопровождающееся уменьшением количества нейронов стриатума и прежде всего синтезирующих ГАМК — стриопаллидарных и стрионигральных нейронов, а также холинергических клеток базальных ганглиев. Отсутствие стрионигрального торможения приводит к гиперактивности дофаминергических клеток и возникновению характерных для хореи непроизвольных судорожных подергиваний.
3.7.7.1. Моделирование синдромов хвостатого ядра
При введении столбнячного токсина, нарушающего, как известно, все виды торможения в ростральную часть хвостатых ядер крыс (Крыжановский Г. Н., Алиев М. Н., 1976) получен комплекс синдромов, включающий стереотипное поведение, кататонию, паркинсонический синдром, ротационный синдром и миоклонию. Синдромы возникали в определенной последовательности и с определенным латентным периодом. Первые признаки стереотипного поведения проявлялись в зависимости от дозы СТ, индивидуальной чувствительности животных и условий опыта (место микроинъекции и др.)- Через 12—18 ч после введения СТ начальные проявления кататонии, которую можно было охарактеризовать отчасти и как каталепсию в связи с повышением пластического тонуса, возникали на фоне стереотипии. На 2—4-й день у животных формировался полный кататонический статус, переходящий затем у некоторых крыс в паркинсонический синдром. Отдельные элементы последнего на-
259
блюдались еще в период развития кататонии, основные же признаки паркинсонизма возникали, как правило, после кататонии. У некоторых животных появлялись миоклонии и джерки с латентным периодом 22—24 ч после введения СТ. Ротационный синдром возникал после введения СТ в одно из хвостатых ядер.
3.7.8. Лимбическая система
Под лимбической системой понимают морфофункциональное объединение, которое включает в себя филогенетически старые отделы коры переднего мозга, а также ряд подкорковых структур, которые регулируют функции внутренних органов, обусловливающих эмоциональную окраску поведения и его соответствие имеющемуся субъективному опыту .
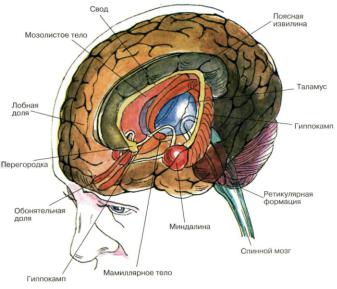
В состав лимбической коры входит древняя кора (палеокортекс), формирующая обонятельный мозг и состоящая из обонятельных луковиц, обонятельного бугорка, прозрачной перегородки и прилежащих областей коры (препериформная, периамигдалярная и диагональная области). Следующий компонент лимбической системы - старая кора (архикортекс), объединяющая сместившийся в процессе эволюции к височной доле гиппокамп (аммонов рог), зубчатую фасцию, основание гиппокампа (субикулум) и расположенную над мозолистым телом поясную извилину
Древняя и старая кора, которые обозначают как аллокортекс, граничат с пятислойной межуточной корой, или мезокортексом, переходящим непосредственно в новую шестислойную кору - неокортекс, или изокортекс. Мезокортекс формируется прилежащей к древней коре островковой, или инсулярной, зоной и граничащей со старой корой парагиппокамповой извилиной (энториальная область и предоснование гиппокампа, или пресубикулум), которые также включены в состав лимбической системы (рис. 58).
Из подкорковых структур в лимбическую систему входят расположенный в медиальной стенке височной доли миндалевидный комплекс и ядра мозговой перегородки. Многие иссле-
260
дователи причисляют к лимбической системе также переднее таламическое ядро, мамиллярные тела и гипоталамус.
Рис. 58. Важнейшие части мозга, образующие лимбическую систему (По Блуму и др.). Атлас «Нервная система человека. Строение и нарушения». Под редакцией В.М.Астапова и Ю.В. Микадзе. 2004.
Все многочисленные формирования лимбической коры кольцеобразно охватывают основание переднего мозга и являются своеобразной границей между новой корой и стволовой частью мозга.
Особенностью лимбической системы является то, что между ее структурами имеются простые двусторонние связи и сложные пути, образующие множество замкнутых кругов. Такая организация создает условия для длительного циркулирования одного и того же возбуждения в системе и тем самым для сохранения в ней единого состояния и навязывание этого состояния другим системам мозга.
В настоящее время хорошо известны связи между структурами мозга, организующие круги, имеющие свою функциональную специфику. К ним относится круг Пейпеца (гиппокамп и сосцевидные тела - передние ядра таламуса - кора поясной извилины - парагиппокампова извилина - гиппокамп). Этот круг имеет отношение к памяти и процессам обучения.
Другой круг (миндалевидное тело - гипоталамус - мезенцефальные структуры - миндалевидное тело) регулирует агрес-