

естественно
.PDF301
с сотр.(Glick, 1982, 1988). Высказанное С.Гликом предположение о том, что преимущественное вращение в правую или левую сторону вызывается наличием дисбаланса между двумя нигростриарными системами, нашло экспериментальное подтверждение. Оказалось, что содержание дофамина :в правом и левом стриатуме различается на 10-15%. При введении амфетамина это различие возрастает на 25%, при чем крысы вращаются преимущественно в направлении, противоположном стороне, содержащей больше дофамина. Первоначально такая асимметрия описывалась как индивидуальная, однако позже у самок было выявлено небольшое, но достоверное предпочтение правосторонних вращательных движений. Эти особи более активны и имеют более выраженную пространственную преференцию. Аналогичный характер асимметрии наблюдался и у других грызунов.
Вращения представляют собой усиленную форму пространственного поведения, и, по-видимому, другие формы пространственной асимметрии также в большей степени связаны с дисбалансом нигростриарной системы. Хорошо известно предпочтение направления движения в лабиринтах различной конструкции. С.И.Глейзер (1981) описал три формы поведения рыб (речных угрей) в х-образном лабиринте. Рыбы с асимметричными формами поведения составляли около половины выборке, причем "правшей" было в среднем в два раза больше, чем "левшей". Индивидуальная пространственно-моторная асимметрия описана у крыс при обучении в лабиринте (Рябинская, 1982 Рябинская, Валуйская, 1983; Hoffman, 1980).
В.В.Тхоржевский (1973) наблюдал преимущественно правостороннее предпочтение у большой выборки мышей. С.Глик и Д.Росс показали, что у крыс дофамина больше в стриатуме, контралатеральном предпочитаемой стороне движения в Т- образном лабиринте. Подчеркивается, что степень выраженности одностороннего предпочтения связана со способностью к обучению. Полагают, что для крыс характерна врожденная, независимая от обучения тенденция .к латерализации, но эта характеристика неустойчива и может полностью модифицироваться при тренировке. Динамические сдвиги пространственномоторной асимметрии наблюдаются при обучении крыс в ла-
302
биринтах разных конструкций (Рябинская, Валуйская, 1983), а также при выработке односторонних побежек в условиях альтернативного выбора зрительных стимулов.
Особое значение имеют результаты специальных исследований роли правого и левого полушарий головного мозга в формировании пространственно-моторного предпочтения. В экспериментах на крысах было установлено различное значение корковых отделов полушарий для формирования простран- ственно-моторного предпочтения в разных экспериментальных условиях: при обучении в лабиринте, при альтернативном выборе зрительных стимулов, а также в тесте "открытое поле".
Для понимания механизмов возникновения асимметрии направления движения существенное значение имеют исследования эффектов стимуляции различных отделов стриарной системы.
Подчеркнем, что специальные исследования различия пространственной асимметрии проводятся в настоящее время в основном на позвоночных животных, причем лишь на немногих и их представителях (рыбы, грызуны, кошки). В эволюционном аспекте перспективным представляется, однако, исследование пространственно-моторной асимметрии и ее мозговых коррелятов у беспозвоночных. При этом следует млеть в виду, что изучение пространственного предпочтения представляет интерес не только в связи с исследованием ФМПА, но и для разработки проблемы пространственной ориентации животных.
Вопрос о латерализации функций у беспозвоночных был поставлен в 1977 г. П.Виттом.. Однако, изучая плетение пауком паутины, он не получил в этом плане положительных результатов. Вместе с тем в литературе можно найти отдельные указания на наличие у беспозвоночных асимметрии направления движения. Так, обнаружено (Wilson, 1968), что в отсутствие зрительной обратной связи летучая саранча может передвигаться либо направо, либо налево, что обусловлено, повидимому, асимметрией моторного контроля. При свободном полете или при наличии симулированного горизонта врожденное асимметричное поведение коррелируется компенсаторными изменениями в моторном паттерне.
Г.А.Мазохин-Цоршняков и В.М.Карцев (1979) показали,

303
что пчелы, обследуя группу близко расположенных объектов, предпочитают двигаться преимущественно против часовой стрелки. У шмелей изучение выбора направления полета при поисках пищи показало стойкое, не зависящее от внешних факторов, левоили правостороннее предпочтение. При этом "правши" и "левши" составляли примерно равное количество. Автор считает, что наблюдавшаяся асимметрия вызвана неточностью развития, а не эволюционной адаптацией, поскольку сильно латерализованные особи не обладали более оптимальной стратегией полета по сравнению с другими особями.
В.Л.Бианки и И.М.Шейман (1985), изучая поведение мучного хрущака в Т-образном; лабиринте, показали, что исходные право и левосторонние поворотные тенденции выражены примерно одинаково. Оказалось, однако, что прочность правосторонних поворотных тенденций более выражена, чем левосторонних, что проявилось как в условиях свободного выбора, так и при переделке исходного предпочтения на противоположное. Пространственно-моторная асимметрия описана также и у ракообразных. Обнаружено достоверное правостороннее предпочтение в 8-лучевом лабиринте и на специальных полигонах у речного рака (Баранюк и др. ,1983). Правостороннее предпочтение наблюдалось и у крабов в экспериментах с активным избеганием. В целом имеющийся литературный материал дает возможность предполагать у насекомых и ракообразных наличие асимметрии направления движения.
В ряде исследований, посвященных изучению латерализации, в качестве свидетельства латерализации использовали предпочтение использования лапы. При исследовании у кошек, обезьян и мышей было выявлено равное предпочтение как левой, так и правой лапой. Эти результаты дали основание предположить, что предпочтение лапы является результатом действия случайных факторов (Спрингер С., Дейч Г., 1983).
К настоящему времени получены многочисленные подтверждения наличия асимметрии функций контроля вокализации, манипуляции, пространственной ориентации и локомоции у различных представителей грызунов, приматов и птиц (В.Л.Бианки, 1985). В частности, показано индивидуальное предпочтение направления движения в открытом поле, в лаби-
304
ринте и в использовании передних конечностей для доставания корма у крыс (В.Л.Бианки 1989).
На различных представителях отряда грызунов-белых крысах, мышах и монгольских песчанках показана моторная асимметрия при развитии индуцированных аудиогенным стрессом состояний тревоги, настороженности, испуга и страха
сисходной дирекциональной ориентацией, последующим круговым бегом, падением на бок, конвульсиями и каталепсией.
Установлено генетически обусловленное, но модифицируемое под воздействием половых стероидов в постнатальный период доминирование правосторонних ротаций у самцов и левосторонних у самок, обусловленное, соответственно, преимущественным возбуждением нейронных систем правого и левого полушарий, что подтверждается электрофизиологическими данными, а также экспериментами с распространяющейся депрессией и воздействием тестостерона и эстрадиола.
Выявленные признаки функциональной асимметрии мозга формируются в раннем возрасте и сохраняются в течение жизни у представителей разных родов и линий, что свидетельствует об их универсальном видоспецифическом характере.
Индекс латерализации сохраняет постоянство при аудиогенном киндлинге и может инвертироваться при одностороннем воздействии блокатора ДА 6-ОДА или РД в контралатеральном полушарии, а также при ограничении звуковосприятия
сипсилатерального уха.
Особый интерес представляли исследования асимметрии мозга у птиц, опубликованные в 1966 год Ф. Ноттебом и его коллегами. Они продемонстрировали, что перерезка у взрослых зябликов и канареек левого подъязычного нерва, который управляет работой мышц левой половины гортани, до неузнаваемости изменяет их пение. Но перерезка правого подъязычного нерва, управляющего правой половиной гортани, влечет минимальные изменения в характере их пения. Это были первые сведения о том, что мозг у птиц обладает выраженной функциональной (левой) асимметрией, что и было подтверждено позднее.
В исследованиях М Петерсена с коллегами (1978), проведённых на японских макаках, обнаружена левосторонняя

305
асимметричность их полушарий по восприятию звуковых сигналов. Обезьян обучали различать два типа звуков, издаваемых особями их вида и определённым/образом реагировать на них. Звуки предварительно записывались на магнитофон и предъявлялись в правое или левое ухо животного в случайном порядке. Обнаружилось, что макаки более точно осуществляли требуемые реакции при трансляции сигнала на правое ухо. По утверждению В. Л. Бианки, имеются многочисленные данные о дви- гательно-пространственных асимметриях даже у беспозвоночных животных: планарий, мучнистого хрущака, речных раков, муравьев и других. (Бианки В. Л., 1989)
Нашими исследованиями (Лобасюк Б.А., 2004) выявлено, что при определении функциональной межполушарной асимметрии (ФМПА) по показателям амплитуд ритмов электрокортикограммы у крыс в разных группах четкой латерализации выявлено не было (рис. 67). Однако при использовании связей-
8
6
4
2
% 0
-2
-4
-6
-8
группа 1 |
группа 2 |
группа 3 |
показатели

 B2A
B2A  B1A
B1A 
 AA
AA 
 TA
TA  DA
DA
Рис. 67. Показатели межполушарной асимметрии ритмов ЭКоГ у крыс 3 групп опытов.
Обозначения: B2A – бета-2 ритм, B1A –бета-1 ритм, AA –альфа ритм, TA –тета ритм, DA – дельта ритм.

306
отношений выявляемых между амплитудами ритмов ЭКоГ с помощью множественной линейной регрессии и корреляции между показателями ЭКоГ четко выявляется различие между правым и левым полушариями головного мозга крыс, во всех трех группах опытов (рис. 68). Поэтому является вполне логичным рассматривать ФМПА как системно организованный феномен.
Большие полушария головного мозга высших позвоночных построены по принципу двусторонней симметрии, т.е. в каждом из них одинаковые структуры расположены в одних и тех же точках полушарий.
АБ
ВГ
ДЕ
Рис. 68. Полициклические мультиграфы отражающие взаимные ориентированные влияния между показателями амплитуд ритмов ЭКоГ Обозначения: А и Б – левое и правое полушария мозга крыс групы №1, В и Г - левое и правое полушария мозга крыс групы №2, , Д и Е - левое и правое полушария мозга крыс групы №3. Узлы графов: 1 - бета-2 ритм, 2 - бета-1 ритм, 3 - альфа ритм, 4 -тета ритм, 5 -дельта ритм. Сплошные линии позитивные влияния, прерывистые – негативные влияния.
307
4.2. Морфологические асимметрии
Взаимодействие гемисфер, необходимое для парной работы, осуществляется посредством межполушарных комиссур: мозолистого тела, передней, задней, габенулярной, гиппокампальной комиссур и промежуточного вещества.
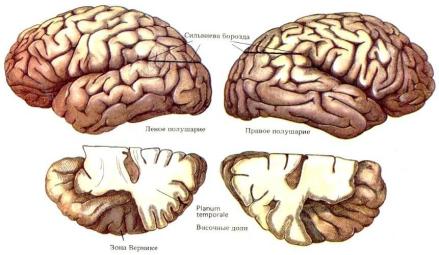
Анатомические исследования височной доли у человека,
шимпанзе и макаки-резуса (Yeni-Komshian G.H. Benson D.A.,
1976), выявили большую длину сильвиевой борозды в височной доле в левом полушарии (рис. 69).
В 1968 году в результате детального посмертного исследования мозга у 100 человек Норман Гешвинд и Уолтер Левитски сообщили о заметных анатомических различиях между полушариями. У 65 из 100 измеренных посмертно экземпляров мозга, участок коры височной доли, перекрывающийся с зоной Вернике и называемый planum temporale, был больше в левом полушарии и у 11 - в правом полушарии. У 24 исследованных экзепляров мозгов случаев различий между полушариями по этому признаку обнаружено не было. В среднем левая височная область была на одну треть была длиннее правой (Gechwind N., Lewitsky, 1968) .
При исследовании 207 экземпляров мозга плодов и новорожденных в возрасте от 10 до 44 недель после зачатия в 54% обнаружили более длинную височную область слева, в 18% справа, а у 28% различий в величине височной плоскости не наблюдалось.
В последующие годы была изучена и измерена не одна сотня других препаратов мозга. Полученные данные в основном совпадают; с данными Гешвинда и Левицкого - приблизительно в 70% случаев planum temporale в левом полушарии крупнее.
Боголеповой И.Н. и ее сотрудниками в макроскопических исследованиях было изучено 205 экземпляров мозга. Было выявлено, что в 54% случаев длина левого полушария превышала длину правого. Однако при исследовании архитектоники полей лобной области 8, 43, 46 и 47 не было выявлено преобладания изученных признаков в одном из полушарий. Авторы предполагают, что это обусловлено сложными интегративными
308
Рис. 69. Анатомическая асимметрия полушарий мозга. Вверху: сильвиева борозда в правом полушарии отклоняется вверх под большим углом. Внизу: задняя часть planum temporale («височная площадка») обычно гораздо больше в левом полушарии, связанном с речевыми функциями (у 70% людей больше в левом, однако у 11% – больше в правом). На эту область частично приходится зона Вернике. (По Гешвинду). Атлас «Нервная система человека. Строение и нарушения». Под редакцией В.М.Астапова и Ю.В. Микадзе. 2004.
функциями лобной области, в которых оба полушария призваны играть важную роль (Боголепова И.Н., Амунц В.В., Оржеховская Н.О., Малофеева Л.И., 1981).
У нормальных людей наиболее четкая асимметрия строения ядер таламуса была выявлена в тех ядрах таламуса, которые тесно связаны с речевыми функциями, в частности, в заднем латеральном ядре таламуса, морфологически связанном с задней височно-теменной областью и нижнетеменной долькой больших полушарий. При двустороннем повреждении заднего латерального ядра зрительного бугра у пациентов возникали явления дизлексии – нарушения чтения (Galaburda A., Eidelberg d. 1982).
Особенности анатомической неравноценности правого и левого полушарий мозга наиболее детально изучены у правшей. Целый ряд структур левого полушария у них характеризуется' большим размером, по сравнению с симметричными отделами правой гемисферы. Так, имеются указания
309
на то, что левая височная доля больше правой на одну треть не только у взрослых (Блинков, Глезер, 1964), но и у детей (Witelson, 1988). При этом особо выраженная (семикратно) нейроанатомическая асимметрия обнаруживается в слуховой коре (Galaburda et al., 1978).
Сходные различия характерны также для задней части постцентральных извилин, обеспечивающих кинестетическую афферентацию артикулярного аппарата (Голдберг и Коста, 1995) и для премоторной области (Galaburda, 1980). Имеются нейроанатомические исследования, указывающие на то, что левый затылочный полюс длиннее и часто выходит за среднюю линию, по сравнению с правым, в то время как правое полушарие шире левого в центральных и фронтальных отделах. Сходные с описанными выше полушарные различия были подтверждены при использовании современных методов исследований, в частности, ядерно-магнитного резонанса
(Charles et al., 1994).
Оценка функциональных различий полушарий у правшей и левшей при выполнении двигательных заданий с помощью современных методов диагностики, в частности, функциональной МРТ, показала большую диффузность реакции у левшей по сравнению с правшами. При этом у левшей, как правило, наблюдалась активация не только контралатеральной, но и соответствующей зоны ипсилатеральной коры.
Имеются также сведения о разнице кровоснабжения полушарий, в частности, о больших размерах внутренней сонной артерии слева и более высоком в ней артериальном давлении (Geshwindt, 1985). Результаты исследования объема мозговых желудочков, согласуются с вышесказанным об асимметрии корковых зон. Сведения о большем объеме левого бокового желудочка и о пропорционально большем количестве серого вещества по отношению к белому веществу в леном полушарии (при большем весе правого полушария), дали основания для заключения о том, что нейроанатомия мола правшей характеризуется выраженной асимметрией. В правом полушарии эти соотношения обратные. При этом кортикальные области, вовлеченные в осуществление речевых сигналов, имеют большую представленность в левом полушарии.
310
Важно отметить, что нейроанатомические различия правшей характерны не только для "речевых" зон, но распространяются и на другие структуры зрительные (Galaburda, 1978), задние ассоциативные (теменные), которые имеют большие размеры В правом полушарии по сравнению с левым. В то же время эти данные позволяют говорить о том, что указанная асимметрия не является модально-специфической -"речевое" против "зрительного" полушария, скорее всего первичные проекционные зоны обеих модальностей наиболее репрезентированы в левой гемисфере мозга. В отличие от этого кортикальные области интермодальных ассоциативных зон более представлены в правом полушарии, в частности, имеются данные о большей величине правого лобного полюса, и о том, что в правом полушарии ассоциативная часть лобной коры, простирающаяся вперед от зоны Брока (Поляков, 1973), представлена в большем объеме. Следует подчеркнуть, что указанные структуры правого полушария вовлечены в наиболее сложные уровни регуляции психической деятельности и принятия решений (Лурия, 1969). При этом они тесно взаимодействуют с задними ассоциативными зонами, образуя единую функциональную систему (Адрианов, 1983).
О нейроанатомической организации мозга левшей имеется меньше сведений, и они свидетельствуют о меньшей асимметрии по сравнению с правшами. Показано, что у 71% левшей расположение сильвиевой борозды симметрично в обоих полушариях. Наряду с этим имеются сведения о больших размерах слуховой коры в правом полушарии по сравнению с левым. Показано также, что у левшей достаточно часто встречаются большие размеры внутренней сонной артерии и более высокое давление в правом полушарии, в то время как у правшей - в левом (Geshwindt, 1985). Знакомство с литературой показывает, что нейроанатомическая асимметрия мозга левшей не является зеркальным" отражением той, которую описывают у правшей, и вариантов асимметрии у них значительно больше, чем у правшей.
Данные о нейроанатомической дифференциации полушарий мозга у правшей позволяют предположить, что выявленные особенности создают предпосылки для неравных