

естественно
.PDF241
ализуется через лобно-мостомозжечковый тракт.
Мозжечок за счет своего влияния на сенсомоторную область коры может изменять уровень тактильной, температурной, зрительной чувствительности. Оказалось, что повреждение мозжечка снижает уровень восприятия критической частоты мельканий света (наименьшая частота мельканий, при которой световые стимулы воспринимаются не как отдельные вспышки, а как непрерывный свет).
Удаление мозжечка приводит к ослаблению силы процессов возбуждения и торможения, нарушению баланса между ними, развитию инертности. Выработка двигательных условных рефлексов после удаления мозжечка затрудняется, особенно в случаях формирования локальной, изолированной двигательной реакции. Точно так же замедляется выработка пищевых условных рефлексов, увеличивается скрытый (латентный) период их вызова.
Влияние мозжечка на вегетативные функции. Мозжечок оказывает угнетающее и стимулирующее влияние на работу сердечнососудистой, дыхательной, пищеварительной и других систем организма. В результате двойственного влияния мозжечок стабилизирует, оптимизирует функции систем организма.
Сердечно-сосудистая система реагирует на раздражение мозжечка либо усилением (например, прессорные рефлексы), либо снижением этой реакции. Направленность реакции зависит от фона, на котором она вызывается. При раздражении мозжечка высокое кровяное давление снижается, а исходное низкое — повышается. Раздражение мозжечка на фоне учащенного дыхания (гиперпноэ) снижает частоту дыхания. При этом одностороннее раздражение мозжечка вызывает на своей стороне снижение, а на противоположной — повышение тонуса дыхательных мышц.
Удаление или повреждение мозжечка приводит к уменьшению тонуса мускулатуры кишечника, из-за низкого тонуса нарушается эвакуация содержимого желудка и кишечника. Нарушается также нормальная динамика секреции и всасывания в желудке и кишечнике.
Обменные процессы при повреждении мозжечка идут более интенсивно, гипергликемическая реакция (увеличение
242
количества глюкозы в крови) на введение глюкозы в кровь или на прием ее с пищей возрастает и сохраняется дольше, чем в норме, ухудшается аппетит, наблюдается исхудание, замедляется заживление ран, волокна скелетных мышц подвергаются жировому перерождению.
При повреждении мозжечка нарушается генеративная функция, что проявляется в нарушении последовательности процессов родовой деятельности. При возбуждении или повреждении мозжечка мышечные сокращения, сосудистый тонус, обмен веществ и т. д. реагируют так же, как при активации или повреждении симпатического отдела вегетативной нервной системы.
Таким образом, мозжечок принимает участие в различных видах деятельности организма: моторной, соматической, вегетативной, сенсорной, интегративной и т. д. Однако эти функции мозжечок реализует через другие структуры центральной нервной системы. Мозжечок выполняет функцию оптимизации отношений между различными отделами нервной системы, что реализуется, с одной стороны, активацией отдельных центров, с другой — удержанием этой активности в определенных рамках возбуждения, лабильности и т. д. После частичного повреждения мозжечка могут сохраняться все функции организма, но сами функции, порядок их реализации, количественное соответствие потребностям трофики организма нарушаются.
На протяжении многих лет мозжечок рассматривался как структура, ответственная исключительно за координацию и статику движений. И хотя в клинических исследованиях давно был накоплен материал о связи патологии мозжечка с такими заболеваниями как аутизм, шизофрения, сенильная деменция, из-за отсутствия инструментально-технической базы эти данные оставались на феноменологическом уровне (Schmahmann J. 1991; Snider S.R. 1982). С развитием нейровизуализационных методов исследования, таких как позитронно-эмиссионная томограмма, однофотонная эмиссионная томограмма и высокоразрешающая магнитно-резонансная томограмма, а также с накоплением огромного массива данных в рамках анатомических, лабораторных, клинических исследований была показана роль
243
мозжечка в организации психических функций (Akshoomoff N.A., Courchersne E.A. 1992). Концепция о влиянии мозжечка на когнитивные процессы сформировалась относительно недавно, но уже в 1997 году в США под редакцией Schahmann была опубликована первая в мире монография «Мозжечок и познание», обобщившая имеющиеся на сегодняшний день клинические, анатомические, физиологические и нейровизуализационные данные (Schmahmann J. 1997). Анатомической основой участия мозжечка в психических функциях являются его двусторонние связи с ассоциативными зонами коры преимущественно контрлатеральных полушарий головного мозга и лимбикоретикулярным комплексом. При этом было отмечено изолированное существование путей, идущих к моторной коре и префронтальным отделам мозга, что в свою очередь определяет возможность изолированного возникновения когнитивных и двигательных расстройств при поражении мозжечка. Показано также, что патология мозжечка различного генеза (опухоли, дегенерации, гипоплазия, сосудистые изменения) приводит к широкому спектру нарушений психических функций в виде нарушений планирования, абстрактного мышления, рабочей памяти, дефицита пространственных функций, речи, эмоциональноличностных изменений..
Однако большинство исследований ограничены рассмотрением либо отдельных психических функций и их составляющих, либо изучением клинических синдромов что препятствует возможности нахождения общих механизмов формирования весьма разнообразных и многочисленных когнитивных симптомов. Представляется адекватным для решения этой задачи применение комплексного синдромного анализа к нарушенным психическим функциям. В школе А.Р.Лурия более 20 лет назад Татьяной Владимировной Мельниковой было проведено исследование больных с субтенториальными опухолями (в том числе и с опухолью мозжечка) и получены данные о наличии у них нейропсихологических синдромов по типу лобного и височно- теменно-затылочного (Мельникова Т.В., 1974). При этом была показана взаимосвязь между нейропсихологическими нарушениями и дисфункцией именно этих отделов коры больших полушарий по ЭЭГ. Однако, выявленные изменения объяснялись
244
преходящими сосудистыми нарушениями, связанными с определенной локализацией опухоли. Роль мозжечка в когнитивном дефиците не была означена и не получила развития в дальнейших исследованиях.
3.7.6.1. Роль полушарий мозжечка в механизмах коркового электрогенеза
С целью изучения влияния мозжечка на механизмы коркового электрогенеза сравнивали показатели электрокортикограммы (ЭКоГ) интактных крыс и крыс с аспирацией полушарий мозжечка. В работе использовались белые крысы массой 210–220 г, которым под нембуталовым наркозом (40 мг/кг, внутрибрюшинно) во фронтальные и окципитальные отделы коры головного мозга билатерально имплантировали отводящие электроды. Вакуумную аспирацию полушарий мозжечка (препарат АП) (пять животных) осуществляли у наркотизированных нембуталом (40 мг/кг) животных. Опыты производили на 3 день после аспирации.
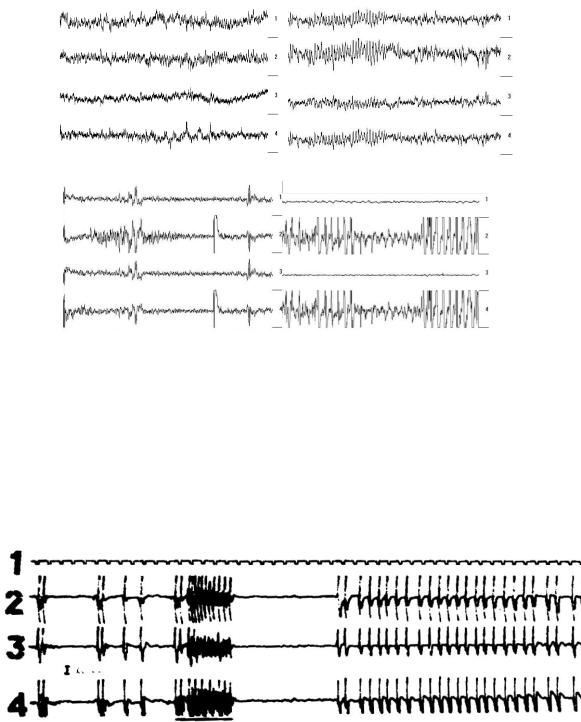
Поскольку все выходы мозжечка ГАМК-эргические, то при аспирации мозжечка в коре головного мозга формируются судорожные потенциалы. На третьи сутки после аспирации полушарий мозжечка у крыс в ЭКоГ выявлялись гиперсинхронные веретена альфа-активности на фоне которых регистрировалась судорожная активность типа пик-волна односторонние отдельные судорожные потенциалы, группы таких потенциалов и билатеральная судорожная активность (рис 54). Следует отметить, что из 5 крыс данной группы опытов у 3 судорожная активность преобладала по амплитуде справа.
При электростимуляции мозжечка ранее сформированная судорожная активность аппликацией конвульсантов (стрихнина и пенициллина) подавлялась (рис. 55).
Анализ фоновой активности ЭКоГ. Сравнение фрагментов синхронизации и десинхронизации крыс с аспирацией полу-
шарий мозжечка. Реорганизация ЭКоГ при переходе от десинхронизации к синхронизации у крыс с аспирацией полушарий мозжечка сопровождалась статистически значимым увеличением всех амплитуд ритмов ЭКоГ в левом и правом полушариях,
245
увеличением частот бета-2 и тета ритмов в обеих полушариях, и частот бета-1 и альфа ритма в левом полушарии. Индексы длительности бета-2 и альфа ритмов ЭКоГ при синхронизации,
А |
Б |
В |
Г |
Рис. 54. ЭКоГ крыс после аспирации полушарий мозжечка. Обозначения: А – интактная крыса, Б-Г – аспирация полушарий мозжечка; 1 – фронтальная кора – затылочная кора слева; 2 – фронтальная кора – затылочная кора справа; 3 – фронтальная кора слева – фронтальная кора справа; 4 – затылочная кора слева – затылочная кора справа. Калибровка для А и Б - 409,6 мкВ, для В и Г - 2048 мкВ, отметка времени 2 сек.
Рис. 55. Влияние электрической стимуляции каудальной части червя мозжечка на эпилептический комплекс (Б) созданные в коре головного мозга кошки аппликацией стрихнина.
Обозначения: 1 – отметка времени, 2- средняя сигмовидная извилин, 3-задняя сигмовидная извилина, 4-передняя сиг-
246
мовидная извилина.
по сравнению с десинхронизацией определялись большими в обеих полушариях. Индекс длительности тета ритма определялся большим, а дельта ритма – меньшим, но только в правом полушарии.
Сравнение фрагментов десинхронизации ЭКоГ у крыс с аспирацией полушарий мозжечка с интактными животными.
У крыс АП по сравнению с ИМ в период десинхронизации в левом полушарии амплитуды бета-2, бета-1, альфа и тета ритмов определялись статистически значимо меньшими. В правом полушарии меньшей определялась только амплитуда бета-2 ритма, в то время как амплитуды бета-1, альфа, тета и дельта ритмов определялись большими. Частота генерации бета-2, альфа и тета ритмов ЭКоГ определялась меньшей, а бета-1 ритма большей, чем у интактных крыс в обеих полушариях. Частота дельта ритма определялась меньшей, только в правом полушарии. Индексы длительности бета-2 и тета ритма у крыс с аспирацией полушарий мозжечка определялись меньшими, а бета-1 и дельта ритма большими, чем у интактных крыс в обеих полушариях.
Сравнение фрагментов синхронизации у крыс с аспирацией полушарий мозжечка с интактными животными. Ам-
плитуда бета-1, альфа, тета и дельта ритмов фрагментов синхронизации у крыс АП определялась статистически большей в обеих полушариях, чем у ИМ. Увеличение амплитуды бета-2 ритма определялось большим только в правом полушарии. Частота бета-1 ритма определялось большей в обеих полушариях, а частоты бета-1 и тета ритмов определялись большими только в левом полушарии. Частоты альфа и дельта ритмов ЭКоГ определялись статистически значимо меньшими только в правом полушарии. Индексы длительности бета-1 ритма ЭКоГ у крыс АП определялись большими в обеих полушариях, а альфа и тета ритма только в левом полушарии. Индекс длительности дельта ритма определялся статистически значимо меньшим только в левом полушарии.
ФМПА показателей амплитуды ЭКоГ у крыс с аспи-
рацией мозжечка. Как было ранее показано (Лобасюк Б.А.,
247
2005), переход паттерна ЭКоГ от десинхронизации к синхронизации в условиях интактности мозга сопровождался сменой левосторонней латерализации амплитуд бета-2-, бета-1-, альфа- и тета-ритмов правосторонней и возрастанием левосторонней латерализации амплитуды дельта-ритма.
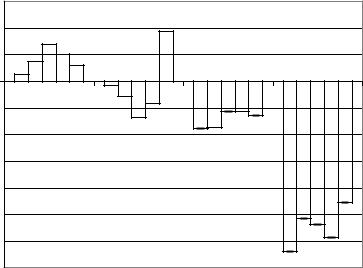
К третьому дню после аспирации полушарий мозжечка ФМПА амплитуды всех ритмов ЭКоГ, (рис. 56) как в условиях десинхронизации, так и в условиях синхронизации выражалась отрицательными величинами, т.е. амплитуды были латерализованы в правое полушарие. В условиях десинхронизации ФМПА амплитуд ритмов ЭКоГ находилась в пределах от (-5,60±0,04%)
15
%
10
5
0
-5
-10
-15
-20
-25
-30
-35
int_desin |
int_sin |
asp_desin |
asp_sin |
|
группы опытов-ритмы ЭКоГ |
|
|

 B2A
B2A  B1A
B1A 
 AA
AA  TA
TA 
 DA
DA
Рис. 56. Диаграммы коэффициентов функциональной межполушарной асимметрии амплитуд ритмов 2, 1, и ЭКоГ, отводимой от интактного мозга и препарата с удаленными полушариями мозжечка.
int_desin – интактный десинхронизация, int_sin – интактный синхронизация, asp_desin – аспирация полушарий мозжечка, десинхронизция, asp_sin – аспирация полушарий мозжечка, синхронизция.
до ( - 8,88±0,03%). При переходе от десинхронизации к синхронизации, латерализация амплитуд ритмов ЭКоГ в правое
248
полушарие возросла, и показатели ФМПА амплитуд ритмов ЭКоГ выражались величинами от (-22,69±0,07%) до (- 31,95±0,05%).
Укрыс с аспирацией полушарий мозжечка по сравнению
синтактными крысами в период десинхронизации в левом полушарии амплитуды бета-2, бета-1, альфа и тета ритмов определялись статистически значимо меньшими. В правом полушарии меньшей определялась только амплитуда бета-2 ритма, в то время как амплитуды бета-1, альфа, тета и дельта ритмов определялись большими. Амплитуда бета-1, альфа, тета и дельта ритмов фрагментов синхронизации у крыс АП определялась статистически значимо большей в обеих полушариях, чем у ИМ.
Таким образом, при аспирации полушарий мозжечка в период десинхронизации в левом полушарии амплитуда определялась меньшей, чем у интактных животных, а в правом – большей, а при синхронизации - большей в обеих полушариях.
Можно предположить, что активирующее (десинхронизирующее) влияние собственно ретикулярной формации ствола мозга ориентировано преимущественно к левому полушарию. Кора левого полушария имеет более развитые связи со стволовыми структурами, тогда как правого – с диэнцефальными. Показано наличие двусторонних связей между мозжечком и полушариями большого мозга. Они представлены корковомостоцеребелярным путем, оканчивающимся в коре мозжечка и церебеллоталамокортикальным, начинающимся в ядрах мозжечка При этом было отмечено изолированное существование путей, идущих к моторной коре и префронтальным отделам мозга (Akshoomjff N.A., Courchersne E. 1992; Schmahmann J., Pandya D., 1987).
Ранее было показано, что показатели ФМПА препарата ИПМ были положительными, т.е. проявлялась отчетливая латерализация электрической активности в левое полушарие.
Учитывая, что все выходы мозжечка ГАМК-ергические, т.е. тормозные, можно предположить, что мозжечок в условиях интактного мозга оказывает тормозное влияние на ствол, и полушария большого мозга. Можно предположить, что при уда-
249
лении полушарий мозжечка проявилось усиленное активационное влияние (десинхронизирующее) ствола мозга на левое полушарие, что в свою очередь, и обеспечило реципрокное усиление механизмов синхронизации в правом полушарии. Это, в свою очередь, и обеспечило латерализацию амплитуд ритмов ЭКоГ в правое полушарие.
В целом можно предположить, что ствол мозга и мозжечок оказывают на ФМПА содружественно-конкурирующее влияние.
3.7.7.Базальные ганглии
Четыре скопления серого вещества, составляющие базальные ганглии (хвостатое ядро, скорлупа, бледный шар, ограда), разделены волокнами белого вещества: внутренней, наружной и внешней капсулами и мозговой полоской. Эти пучки волокон, отграничивая базальные ганглии друг от друга, связывают их между собой и с соседними структурами сложной системой проводящих путей.
Работы Капперса (Kappers et al., 1960) привели к созданию концепции, согласно которой хвостатое ядро (ХЯ) и скорлупа рассматривались как неостриатум, а бледный шар — как палеостриатум. Бледный шар обычно подразделен на внутренний и наружный сегменты. У хищников и грызунов Внутренний сегмент ясно отграничен и существует как энтопедункулирное ядро. Отмечалось более раннее созревание волокон во внутреннем сегменте бледного шара.
Хвостатые ядра представляют собой весьма сложное в морфологическом и функциональном отношениях образование. Оно связано со многими образованиями мозга и играет роль модулятора деятельности высших отделов ЦНС. Помимо классических представлений о стриатуме как о высшем центре экстрапирамидной системы, все более укрепляется мнение об участии этого отдела ЦНС в механизмах высшей интегративной деятельности мозга, психической деятельности человека и поведения животных. Поэтому патология хвостатых ядер имеет многообразное выражение в зави-
250
симости от того, какие структуры и медиаторные м е- ханизмы ядра вовлечены в патологический процесс.
Следующие медиаторные аппараты являются, по-видимому, основными и функциональной структуре неостриатума.
Дофаминергический аппарат. Нигростриарный пул, состоящий из аксонов нейронов, расположенных в компактной зоне черной субстанции, имеет в основном дофаминергическую природу. В хвостатом ядре дофаминергические терминалы контактируют преимущественно с холинергическими интернейронами II тина (Hattori Т. et al, 1976).
Некоторые терминалы контактируют с нехолинергическими интернейронами, часть из которых является гамкергическими. На крупных холинергических нейронах, которые являются выходными и дают эфферентные волокна, дофаминергические терминалы, по-видимому, не оканчиваются. Дофамин является медиатором торможения в хвостатом ядре, в частности, по отношению к холинергическим нейронам. Агонисты дофамина также вызывают торможение этих нейронов, в то время как антагонисты их активируют. Эффекты дофамина такие же, как и эффекты раздражений черной субстанции. Вместе с тем дофамин может вызывать возбуждение небольшой части нейронов. Предполагается, что нейроны, активируемые дофамином, являются тормозными интернейронами, ограничивающими активность эффекторных нейронов стриатума.
Согласно некоторым данным, в хвостатом ядре имеются интернейроны недофаминергической природы, возбуждение которых опосредуется через возвратные коллатерали эффекторных нейронов. Возможно, что дофамин действует на пресииантические окончания ГАМК-ергических нейронов, что приводит к ограничению выброса ГАМК и растормаживанию последующего ГАМК-рецептивного нейрона. Таким образом, в целом поступающие в стриатум дофаминергические волокна вызывают преимущественно тормозные эффекты: они тормозят активность возбуждающих холинергических нейронов, активируют тормозные (предположительно