

Лекция № 19-21. Химический состав и структура нуклеиновых кислот
1. Химический состав нуклеиновых кислот
Нуклеиновые кислоты — сложные высокомолекулярные соединения, обеспечивающие хранение, передачу наследственной информации и реализацию этой информации. Их структурные компоненты выполняют функции кофакторов, аллостерических эффекторов, входят в состав коферментов, принимают участие в обмене веществ и энергии. При полном гидролизе нуклеиновых кислот образуются пуриновые и пиримидиновые основания, углеводы (рибоза и дезоксирибоза) и фосфорная кислота. Углеводы — рибоза и дезоксирибоза — в молекулах нуклеиновых кислот находятся в e-D-рибофуранозной форме:
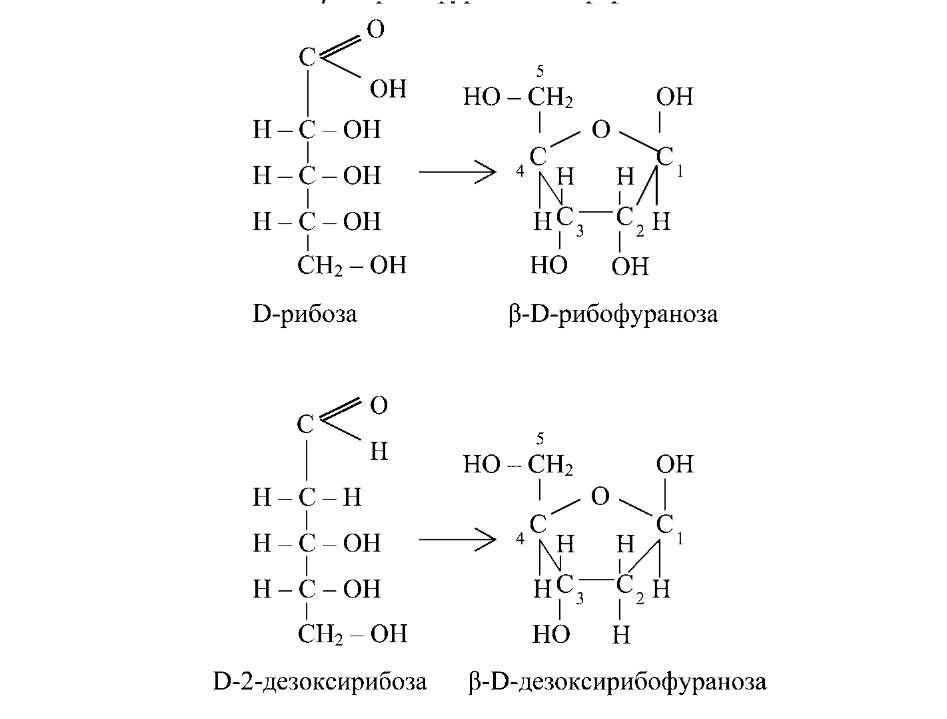
Углевод в молекуле РНК представлен рибозой, а в молекуле ДНК — дезоксирибозой, откуда и их названия. Основой структуры азотистых оснований являются пурин и пиримидин:
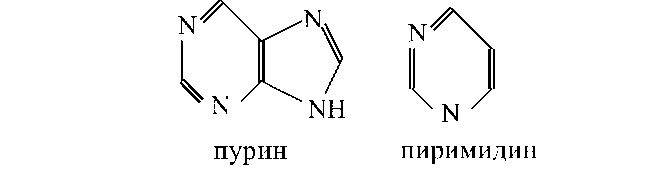
В составе нуклеиновых кислот встречаются три главных пиримидиновых основания: цитозин, урацил и тимин:
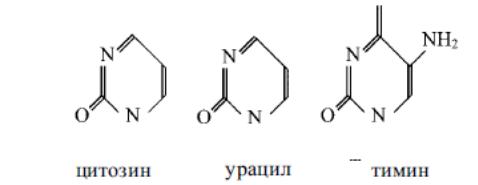
Существуют также минорные пиримидиновые основания: 5-метил и 5-оксиметилцитозин, дигидроурацил, псевдоурацил, 1-метилурацил, оротовая кислота, 5-карбоксиурацил, 4-тиоурацил и др. Главные пуриновые основания, входящие в состав нуклеиновых кислот, — это аденин и гуанин:
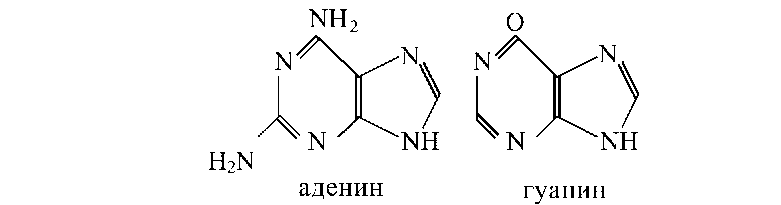
К минорным пуиновым основаниям относятся инозин, N-метиладенозин, N-метилгуанозин, ксантин, гипоксантин, 7-метилгуанозин и др.
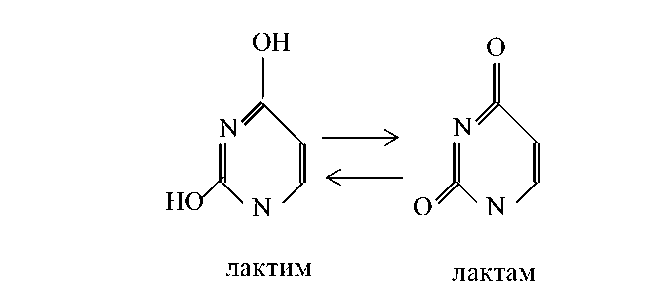
Азотистые основания способны существовать в двух таутомерных формах: лактимной и лактамной (при pH 7,0 они находятся в лактамной форме, при снижении pH переходят в лактимную форму).
Все оксипроизводные природных нуклеиновых кислот находятся в лактамной форме.
2. Особенности структуры нуклеиновых кислот
Существует ряд закономерностей состава и количественного содержания азотистых оснований в нуклеиновых кислотах (правила Чаргаффа).
Молярная доля пуринов равна молярной доле пиримидинов: А + Г = Ц + Т или (А + Г) / (Ц + Т) = 1.
Количество аденина и цитозина равно количеству гуанина и тимина: А + Ц = Г + Т или (А + Ц) / (Г + Т) = 1.
Количество аденина равно количеству тимина, а количество гуанина равно количеству цитозина: А = Т и Г = Ц или А / Т = 1 и Г / Ц = 1.
Коэффициент специфичности (Г + Ц) / (А + Т) для животных и большинства растений ниже 1 (0,54—0,94), у микроорганизмов он может быть выше 1 (0,45—2,57).
Структурными единицами нуклеиновых кислот являются нуклеотиды, состоящие из азотистого основания, углевода и фосфорной кислоты, причем углевод занимает центральное место:

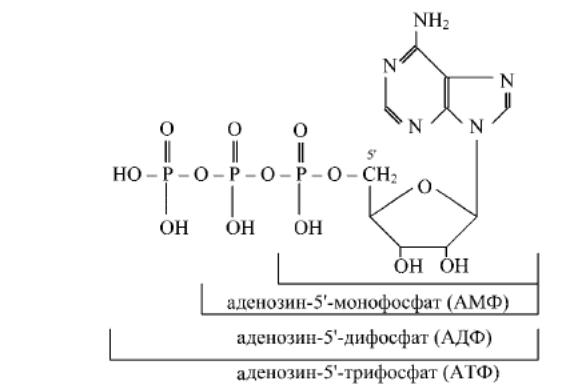
Пентоза соединена с остатком фосфорной кислоты сложноэфирной связью, а с азотистым основанием — N-гликозидной связью. При отщеплении фосфорной кислоты образуются нуклеозиды. В качестве радикала R может выступать H- либо OH-группа. Различают пять типов мононуклеотидов в зависимости от азотистого основания — АМФ, ГМФ, УМФ, ЦМФ и ТМФ. Мононуклеотиды , присоединив к себе еще один остаток фосфорной кислоты, образуют нуклеозиддифосфаты. При присоединении еще одного остатка фосфорной кислоты образуются нуклеозидтрифосфаты. Только нуклеозидтрифосфаты используются для синтеза нуклеиновых кислот.
В организме существуют также циклические нуклеотиды:

Циклический 2', 3'-АМФ образуется в качестве промежуточного продукта распада РНК, а циклический 3', 5'-АМФ встречается в свободном состоянии, играя роль медиатора внеклеточных сигналов у животных. Аналогичная функция у цГМФ, производных УДФ и ЦТФ и нуклеотидов в составе кофакторов и коферментов.
Синтетические аналоги азотистых оснований, нуклеозидов и нуклеотидов используются в медицине (в онкологии). Встраиваясь в клеточные компоненты, они оказывают цитотоксическое действие. К ним относятся 5-фторурацил, 6-тио и 6-меркаптопурин, 8-азагуанин, 6-азауридин и 6-азацитидин, 5-йодпроизводное дезоксиуридина.