

Лекция №47 метаболизм фосфолипидов
В отличие от триглицеридов и жирных кислот фосфолипиды не являются существенным энергетическим материалом. Фосфолипиды играют важную роль в структуре и функции клеточных мембран, активации мембранных и лизосомальных ферментов, в проведении нервных импульсов, свертывании крови, иммунологических реакциях, процессах клеточной пролиферации и регенерации тканей, в переносе электронов в цепи «дыхательных» ферментов. Особая роль фосфолипидам отводится в формировании липо-протеидных комплексов.
Биосинтез фосфолипидов интенсивно происходит в печени, стенке кишечника, семенниках, яичниках, молочной железе и других тканях. Наиболее важные фосфолипиды синтезируются главным образом в эндоплазма-тической сети клетки.
Центральную роль в биосинтезе фосфолипидов играют 1,2-диглицериды (в синтезе фосфатидилхолинов и фосфатидилэтаноламинов), фосфатидная кислота (в синтезе фосфатидилинозитов) и сфингозин (в синтезе сфин-гомиелинов). Цитидинтрифосфат (ЦТФ) участвует в синтезе практически всех фосфолипидов. В качестве примера рассмотрим синтез отдельных представителей фосфолипидов.

Биосинтез фосфатидилэтаноламина. Первоначально этаноламин при участии соответствующей киназы фосфорилируется с образованием фосфо-этаноламина:
Затем фосфоэтаноламин взаимодействует с ЦТФ, в результате чего образуются цитидиндифосфатэтаноламин (ЦДФ-этаноламин) и пирофосфат (PPi):
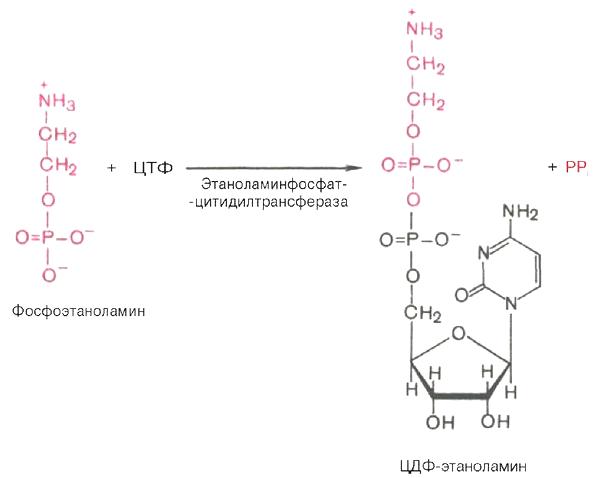
В следующей реакции ЦДФ-этаноламин, взаимодействуя с 1,2-дигли-церидом, образующимся при дефосфорилировании фосфатидной кислоты, превращается в фосфатидилэтаноламин. Реакция катализируется ферментом этаноламинфосфотрансферазой:
ЦДФ-этаноламин + 1,2-диглицерид -> Фосфатидилэтаноламин + ЦМФ.
Биосинтез фосфатидилхолина (лецитина). Фосфатидилэтаноламин является предшественником фосфатидилхолина. В результате последовательного переноса трех метальных групп от трех молекул S-аденозилметионина (донор метальных групп, см. главу 6) к аминогруппе остатка этаноламина образуется фосфатидилхолин:
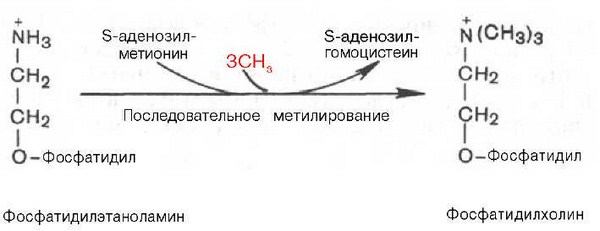
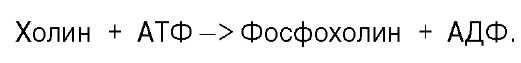
Затем фосфохолин реагирует с ЦТФ, образуя цитидиндифосфатхолин (ЦДФ-холин):
![]()
В дальнейшем ЦДФ-холин взаимодействует с 1,2-диглицеридом, в результате чего образуется фосфатидилхолин:
![]()
Биосинтез фосфатидилсерина. У млекопитающих фосфатидилсерин образуется в реакции обмена этаноламина на серин следующим путем:
![]()
Существует еще один путь синтеза фосфатидилхолина в клетках животных. В этом случае, как и при синтезе фосфатидилэтаноламина, используется ЦТФ в качестве переносчика, но уже не фосфоэтаноламина, а фос-фохолина. На первом этапе синтеза свободный холин активируется под действием холинкиназы с образованием фосфохолина:

![]()
Существует и второй
путь образования фосфатидилсерина,
который связан с предварительным
вовлечением фосфатидной кислоты в
синтез фосфоглицеридов: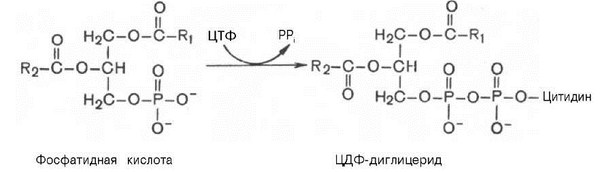
Следует отметить, что различие в синтезе холин – и этаноламинсодер-жащих фосфолипидов, с одной стороны, и инозитсодержащих фосфолипидов —с другой, заключается в том, что в первом случае при участии ЦТФ образуется ЦДФ-холин или ЦДФ-этаноламин—реакционноспособные азотистые основания, а во втором случае при участии ЦТФ образуется ЦДФ-диглицерид — реакционноспособная форма диглицерида.
Распад и обновление фосфолипидов
Известно, что молекулы белков расщепляются в тканях полностью. Поэтому для молекулы белков можно определить время обновления. Фосфоли-пиды также активно распадаются в тканях, но для каждой части молекулы время обновления различно. Например, время обновления фосфатной группы отличается от времени обновления 1-ацильной группы, и обусловлено это наличием ферментов, вызывающих частичный гидролиз фосфолипидов, вслед за которым снова может происходить их синтез (рис. 11.7).

К сожалению, в настоящее время нет достаточно полных данных о фосфолипазном спектре той или иной ткани. Хорошо известно, что фосфолипаза A1 атакует эфирную связь фосфолипидов в положении 1. Фосфолипаза A2 катализирует гидролиз эфирной связи в положении 2 гли-церофосфолипидов, в результате чего образуются свободная жирная кислота и лизофосфолипид (в случае фосфатидилхолина — лизолецитин), который реацилируется ацил-КоА при участии ацилтрансферазы.
Фосфолипаза С атакует эфирную связь в положении 3, что заканчивается образованием 1,2-диглицерида и фосфорильного основания.
Фосфолипаза D катализирует отщепление от фосфолипида азотистого основания. Долгое время считалось, что фосфолипаза D содержится только
В растительных тканях. В последнее время ее удалось обнаружить в растворимой фракции мозга крысы, а затем в микросомах мозга и других органов, а в самое последнее время-в митохондриях печени крысы.
Нет ясности в отношении фосфолипазы В. Возможно, что это-смесь ферментов, обладающих свойствами фосфолипаз А1 и А2. Не исключено, что фосфолипаза В-фермент, действующий только на лизофосфолипид (например, лизолецитин), т. е. это лизофосфолипаза.