

Лекция №56-57 синтез и распад гликогена
Гликоген—главная форма запасания углеводов у животных и человека. Накапливается гликоген главным образом в печени (до 6% от массы печени) и в скелетных мышцах, где его содержание редко превышает 1%. Запасы гликогена в скелетных мышцах ввиду значительно большей массы последних превышают его запасы в печени. Гликоген присутствует в цито-золе в форме гранул диаметром от 10 до 40 нм. На электронных микрофотографиях гликогеновые гранулы выглядят плотными. Установлено, что эти гранулы, кроме гликогена, содержат ферменты, катализирующие синтез и распад гликогена. Однако гликогеновые гранулы отличаются от мульти-ферментных комплексов (например, от пируватдегидрогеназного комплекса). Степень структурной организации гликогеновых гранул ниже, чем в мультиферментных комплексах. Следует подчеркнуть, что синтез и распад гликогена в клетке осуществляются разными метаболическими путями. В частности, считалось, что гликогенфосфорилаза (фосфорилаза а) катализирует как распад гликогена, так и его синтез, потому что в опытах in vitro было показано, что гликогенфосфорилазная реакция обратима. Однако в дальнейшем было установлено, что в клетке (in vivo) фосфорилаза а катализирует только распад гликогена, синтез гликогена осуществляется при участии совершенно другого фермента. Оба эти процесса (синтез и распад гликогена) регулируют содержание глюкозы в крови и создают резерв глюкозы для интенсивной мышечной работы.
Синтез гликогена (гликогенез)
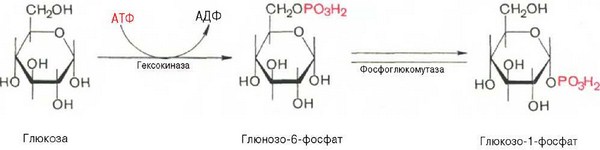
Прежде всего глюкоза подвергается фосфорилированию при участии фермента гексокиназы, а в печени—и глюкокиназы. Далее глюкозо-6-фосфат под влиянием фермента фосфоглюкомутазы переходит в глюкозо-1-фос-фат:
![]()
Приводим структурную формулу УДФ-глюкозы:
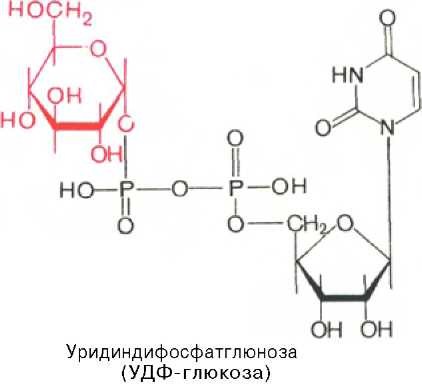
Образовавшийся глюкозо-1-фосфат уже непосредственно вовлекается в синтез гликогена. На первой стадии синтеза глюкозо-1-фосфат вступает во взаимодействие с УТФ (уридинтрифосфат), образуя уридиндифосфатглю-козу (УДФ-глюкоза) и пирофосфат. Данная реакция катализируется ферментом глюкозо-1-фосфат-уридилилтрансферазой (УДФГ-пирофосфорила-за):На второй стадии — стадии образования гликогена — происходит перенос глюкозного остатка, входящего в состав УДФ-глюкозы, на глюкозидную цепь гликогена («затравочное» количество). При этом образуется а-(1—>4)-связь между первым атомом углерода добавляемого остатка глюкозы и 4-гидроксильной группой остатка глюкозы цепи. Эта реакция катализируется ферментом гликогенсинтазой. Необходимо еще раз подчеркнуть, что реакция, катализируемая гликогенсинтазой, возможна только при условии, что полисахаридная цепь уже содержит более 4 остатков D-глюкозы.
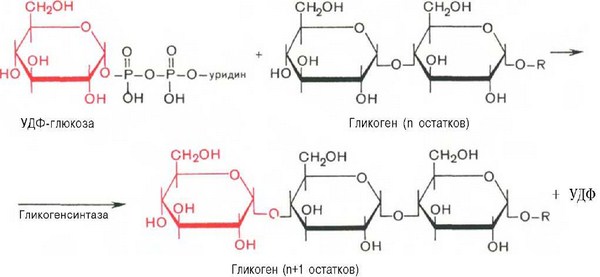
Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ, и таким образом весь цикл превращений глюкозо-1-фосфата начинается сначала.
В целом образование а-1,4-глюкозидной ветви («амилозной» ветви) гликогена можно представить в виде следующей схемы:
Установлено, что гликогенсинтаза неспособна катализировать образование а-(1—>6)-связи, имеющейся в точках ветвления гликогена. Этот процесс катализирует специальный фермент, получивший название гли-когенветвящего фермента, или амило-(1—>4)—>(1—>6)-трансглюкозидазы. Последний катализирует перенос концевого олигосахаридного фрагмента, состоящего из 6 или 7 остатков глюкозы, с нередуцирующего конца одной из боковых цепей, насчитывающей не менее 11 остатков, на 6-гидроксиль-ную группу остатка глюкозы той же или другой цепи гликогена. В результате образуется новая боковая цепь.
Ветвление повышает растворимость гликогена. Кроме того, благодаря ветвлению создается большое количество невосстанавливающих концевых остатков, которые являются местами действия гликогенфосфорилазы и гликогенсинтазы.
Таким образом, ветвление увеличивает скорость синтеза и расщепления гликогена.
Благодаря способности к отложению гликогена (главным образом в печени и мышцах и в меньшей степени в других органах и тканях) создаются условия для накопления в норме некоторого резерва углеводов. При повышении энерготрат в организме в результате возбуждения ЦНС обычно происходят усиление распада гликогена и образование глюкозы.
Помимо непосредственной передачи нервных импульсов к эффекторным органам и тканям, при возбуждении ЦНС повышаются функции ряда желез внутренней секреции (мозговое вещество надпочечников, щитовидная железа, гипофиз и др.), гормоны которых активируют распад гликогена, прежде всего в печени и мышцах (см. главу 8).
Как отмечалось, эффект катехоламинов в значительной мере опосредован действием цАМФ, который активирует протеинкиназы тканей. При участии последних происходит фосфорилирование ряда белков, в том числе гликогенсинтазы и фосфорилазы b — ферментов, участвующих в обмене углеводов. Фосфорилированный фермент гликогенсинтаза сам по себе малоактивен или полностью неактивен, но в значительной мере активируется положительным модулятором глюкозо-6-фосфатом, который увеличивает Vmax фермента. Эта форма гликогенсинтазы называется D-формой, или зависимой (dependent) формой, поскольку ее активность зависит от глюкозо-6-фосфата. Дефосфорилированная форма гликогенсинтазы, называемая также I-формой, или независимой (independent) формой, активна и в отсутствие глюкозо-6-фосфата.
Таким образом, адреналин оказывает двойное действие на обмен углеводов: ингибирует синтез гликогена из УДФ-глюкозы, поскольку для проявления максимальной активности D-формы гликогенсинтазы нужны очень высокие концентрации глюкозо-6-фосфата, и ускоряет распад гликогена, так как способствует образованию активной фосфорилазы а. В целом суммарный результат действия адреналина состоит в ускорении превращения гликогена в глюкозу.
Распад гликогена (гликогенолиз)
Известно, что фосфоролитический распад играет ключевую роль в мобилизации полисахаридов.
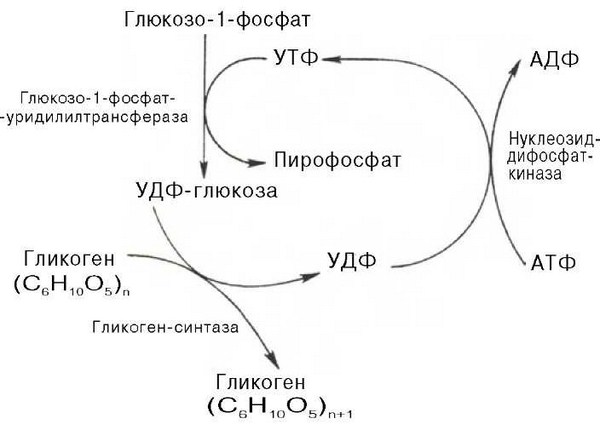
Фосфорилазы переводят полисахариды (в частности, гликоген) из запасной формы в метаболически активную форму; в присутствии фосфорилазы гликоген распадается с образованием фосфорного эфира глюкозы (глюкозо-1-фосфата) без предварительного расщепления на более крупные обломки молекулы полисахарида. В общей форме эту реакцию можно представить в следующем виде:
![]()
Где (С6Н10О5)п означает полисахаридную цепь гликогена, а (С6Н10О5)п,— ту же цепь, но укороченную на один глюкозный остаток.
На рис. 10.1 изображены
процесс распада гликогена до
глюкозо-1-фосфата и участие в этом
процессе цАМФ. Фермент фосфорилаза
существует в двух формах, одна из которых
(фосфорилаза а) активна, в то время как
другая (фосфорилаза b) обычно неактивна.
Обе формы могут диссоциировать на
субъединицы. Фосфорилаза b состоит из
двух субъединиц, а фосфорилаза а—из
четырех. Превращение фосфо-рилазы b в
фосфорилазу а осуществляется
фосфорилированием белка:![]()
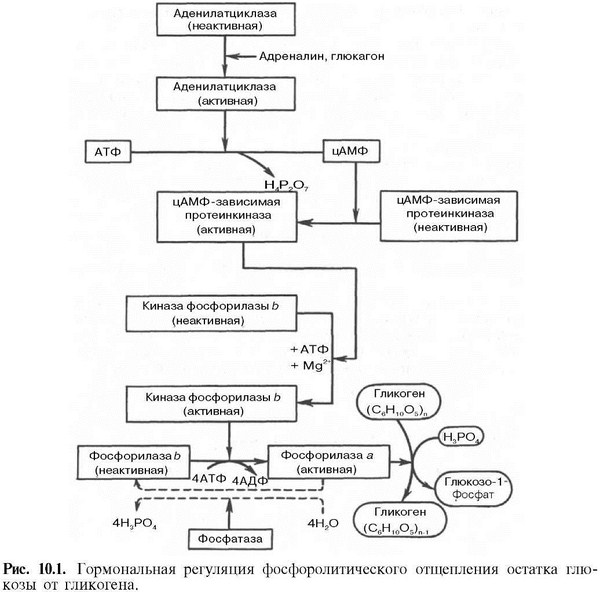
Катализируется эта реакция ферментом, который называется киназой фосфорилазы b. Установлено, что эта киназа может существовать как в активной, так и в неактивной форме. Неактивная киназа фосфорилазы превращается в активную под влиянием фермента протеинкиназы (киназа киназы фосфорилазы), и не просто протеинкиназы, а цАМФ-зависимой протеинкиназы.
Активная форма последней образуется при участии цАМФ, которая в свою очередь образуется из АТФ под действием фермента аденилатцик-лазы, стимулируемой, в частности, адреналином и глюкагоном. Увеличение содержания адреналина в крови приводит в этой сложной цепи реакций к превращению фосфорилазы b в фосфорилазу а и, следовательно, к освобождению глюкозы в виде глюкозо-1-фосфата из запасного полисахарида гликогена. Обратное превращение фосфорилазы а в фосфорилазу b катализируется ферментом фосфатазой (эта реакция практически необратима).
Образовавшийся в результате фосфоролитического распада гликогена глюкозо-1-фосфат превращается под действием фосфоглюкомутазы в глюкозо-6-фосфат. Для осуществления данной реакции необходима фосфо-рилированная форма фосфоглюкомутазы, т. е. ее активная форма, которая образуется, как отмечалось, в присутствии глюкозо-1,6-бисфосфата.

Образование свободной глюкозы из глюкозо-6-фосфата в печени происходит под влиянием глюкозо-6-фосфатазы. Данный фермент катализирует гидролитическое отщепление фосфата:

Заметим, что фосфорилированная глюкоза в противоположность неэте-рифицированной глюкозе не может легко диффундировать из клеток. Печень содержит гидролитический фермент глюкозо-6-фосфатазу, который и обеспечивает возможность быстрого выхода глюкозы из этого органа. В мышечной ткани глюкозо-6-фосфатаза практически отсутствует.
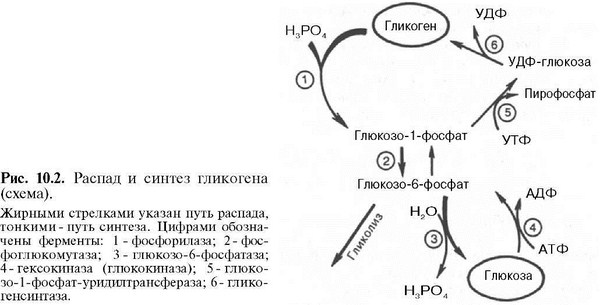
На рис. 10.2 отражены представления о путях распада и синтеза гликогена в печени.
Можно считать, что сохранение постоянства концентрации глюкозы в крови является результатом одновременного протекания двух процессов: поступления глюкозы в кровь из печени и потребления ее из крови тканями, где она используется в первую очередь как энергетический материал.
В тканях (в том числе в печени) распад глюкозы происходит двумя основными путями: анаэробным (при отсутствии кислорода) и аэробным, для осуществления которого необходим кислород.