

Лекция № 22. Обмен нуклеиновых кислот
1. Общие представления об обмене нуклеопротеидов
Нуклеиновые кислоты имеют два источника: поступление с пищей и саморепродукция. Распад нуклеотидов (соединений, входящих в состав нуклеиновых кислот) дает мочевую кислоту, которая как конечный продукт пуринового обмена выделяется с мочой. Переваривание нуклеопротеидов начинается в желудке. Под влиянием соляной кислоты и пепсина разрушаются связи между белковым компонентом нуклеопротеидов и их простетической группой. Нуклеиновые кислоты в кишечнике гидролизуются под действием ДНКазы и РНКазы панкреатического сока, продукты гидролиза — олиго- и мононуклеотиды. Фосфатазы в кишечнике гидролизуют мононуклеотиды до нуклеозидов и фосфорной кислоты. В тканях нуклеотиды служат предшественниками и составными элементами нуклеиновых кислот, участвуют в кумуляции энергии и ее транспорте (АДФ-АТФ), являются структурными компонентами коферментов (НАД, НАДФ, ФАД, КоА, ГТФ, ЦТФ), посредниками при передаче гормональных сигналов (3, 5-цАМФ, 3,5-цГМФ) на внутриклеточные системы.
Биосинтез пуриновых и пиримидиновых нуклеотидов осуществляется клетками большинства тканей. Пуриновый скелет образуется из фрагментов разных соединений: аспартата фор-мила, глутамина, углекислоты и глицина.
Синтез начинается с образования 5-фосфорибозил-1-амина, далее к аминогруппе присоединяется остаток глицина. Затем в образовании пуринового ядра участвуют метенил-Н4-фолат, глутамин, диоксид углерода, аспарагиновая кислота, формильный остаток формил-Н4-фолата.
Эта совокупность реакций ведет к образованию инозиновой кислоты (ИМФ), которая является предшественником АМФ и ГМФ. Специфические киназы превращают нуклеозидмонофосфаты в нуклеозидди- и трифосфаты. Высвобождающиеся в процессе превращения нуклеотидов в тканях пуриновые основания (аденин и гуанин) могут использоваться повторно для синтеза нуклеотидов.
Пиримидиновое ядро пиримидиновых нуклеотидов синтезируется из диоксида углерода, амидной группы глутамина и аспарагиновой кислоты.
Первый этап синтеза — образование карбамоилфосфата, поставщиком аминогруппы для которого является глутаминовая кислота, затем из него образуются карбамоиласпартат, дигидрооротат, оротат, оротидиловая кислота, уридиловая кислота (УМФ).
Цитидиловые нуклеотиды образуются по следующему пути:
У МФ
+ АТФ УДФ +АДФ
МФ
+ АТФ УДФ +АДФ
У ДФ + АТФ УТФ + АДФ.
УТФ затем аминируется в ЦДФ за счет аминогруппы глутамина. Тимидиловые нуклеотиды происходят из дезоксиуридидиловой кислоты (дУМФ), реакция катализируется тимидилатсинтетазой, донор одноуглеродистого фрагмента — метенил-Н4-фолат.
Дезоксирибонуклеотиды образуются из рибонуклеотидов за счет восстановления остатка рибозы: донором водорода служит низкомолекулярный белок тиоредоксин, содержащий две тиогруппы. За счет водорода этих групп кислород рибозы в положении С-2 восстанавливается до молекулы воды, а другой фермент (тиредоксинредуктаза) восстанавливает окисленный тиоредоксин за счет водорода НАДФ*Н.
Пути образования дезоксирибонуклеотидов:
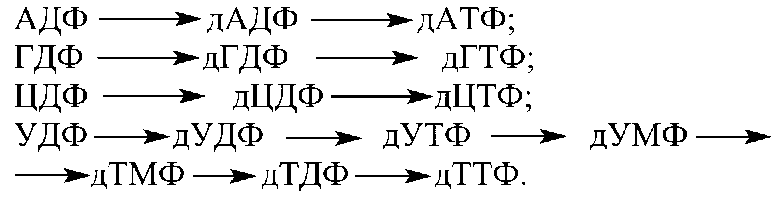
Активация синтеза дезоксирибонуклеотидов наблюдается на стадиях, предшествующих делению клетки. Катаболизм пуриновых нуклеотидов завершается образованием мочевой кислоты. На первом этапе АМФ, теряя гидролитически рибозофосфат и аминогруппу, превращается в гипоксантин, затем в ксантин. ГМФ, теряя рибозофосфатный остаток и аминогруппу, также превращается в ксантин, окисление последнего приводит к образованию мочевой кислоты, которое катализирует ксантиоксидаза. Катаболизм пиримидиновых нуклеотидов. Цитозин в результате дезаминирования превращается в урацил. Затем после гидролитического раскрытия пиримидинового кольца, дезаминирования и декарбоксилирования в качестве конечного продукта образуется β-аланин.
Тимин подвергается тем же превращениям, что и урацил, конечный продукт его превращений — β-аминомасляная кислота. Нарушение обмена пуриновых и пиримидиновых оснований: основной конечный продукт пуринового обмена — мочевая кислота — транспортируется кровью в комплексе с уратсвязывающим протеином и полностью фильтруется в клубочке. Нарушения обмена мочевой кислоты могут быть связаны с ее избыточным образованием, снижением содержания уратсвязывающего протеина или нарушением выделения почками. Классическим проявлением патологии является подагра, это результат генетически обусловленных энзимодефектов.